Анатомия опорно двигательная система. Информационно-дидактический блок (1). Закономерная последовательность смены периодов клеточного цикла
 Скачать 426.5 Kb. Скачать 426.5 Kb.
|

|
Информационно- дидактический блок Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков, как циклины и циклин-зависимые киназы (Ц-ЦЗК). Присоединение циклина не только активирует ЦЗК, но и обеспечивает ее специфичное действие по отношению к тем или иным белкам. Концентрация циклина в клетке в течение клеточного цикла изменяется циклическим образом. Клеточный цикл контролируется изменением активности Cdk, которая регулируется периодическим образованием и распадом их регуляторных субъединиц – циклинов. Смена синтезов и разрушений различных циклинов обеспечивает переходы и протекания различных фаз клеточного цикла. В разные фазы клеточного цикла образуются разные циклины, которые связываясь с Cdk образуют Cdk-циклиновые комплексы (ц-Cdk). В этом комплексе циклин является активаторной, а ЦЗК – каталитической субъединицей (рис.1). Таким образом, контроль регуляции митотического цикла происходит в результате 1.присоединения циклина к циклин-зависимой киназе (Cdk) 2.фосфорилирование и дефосфорилирование Cdk 3.ингибирование специальными белками семейства INK4 и KIP1. И так в основе механизма действия комплексов Ц-ЦЗК лежат общие последовательные процессы: а) инактивация регулирующих факторов предыдущей стадии митотического цикла; б) активизация факторов, обеспечивающих данную стадию митотического цикла; в) образование и активизация комплекса факторов, необходимых для следующей стадии деления клетки. 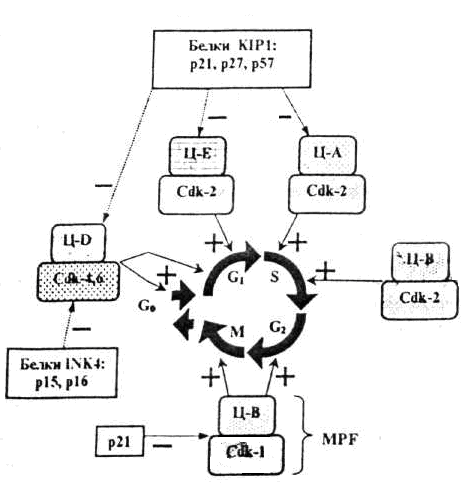 Рис. 1. Комплексы циклин- Cdk, определяющие разные фазы клеточного цикла Ц- циклины, Cdk – циклинзависимые киназы, MPF –митозстимулирующий фактор. В прямоугольных рамках – ингибиторы комплексов. Для эффективного начала деления клетки необходимы не только благоприятные внешние условия, но и специальные сигнальные молекулы – ростовые факторы (митогены)и прикрепление клетки к какой-либо поверхности (эпителиоциты – к базальной мембране, фибробласты – к коллагеновым волокнам), которые связываются с рецепторами на поверхности клеток-мишеней, так же отсутствие контактов с другими клетками запускают механизм митотического цикла. Сигнальные пути регуляции клеточного цикла Инициация процессов митотического цикла происходит под влиянием комплекса циклин Д-ЦЗК-4 и/или циклина Д-ЦЗК-6. Для активации инициирующего комплекса необходимо поступление сигналов из внеклеточных регуляторов – фактора роста, т.е. эпидермального фактора роста (ЭФР) и антигенпредставляющей клетки (АПК). Сигнал, воспринимаемый рецепторными тирозинкиназами, активизирует каскадным путем киназный домен рецептора, митогенактивируемые протеинкиназы (МАПК), которые являются транскрипционными факторами (ТФ) для генов раннего ответа (FOS, JUN), быстро отвечающих на воздействие митогена. Продукты генов раннего ответа (FOS, JUN) в свою очередь становятся ТФ для генов замедленного ответа и активируют их. Продукты генов замедленного ответа, в т.ч. белки-ферменты, циклин Д-ЦЗК 4, ЦЗК 6, приводят к образованию комплексов циклин–ЦЗК, действующих в первой половине G1-периода митотического цикла. Кроме того, активируются и другие гены, в т.ч. ген белка Мус. Белок Мус тормозит активность гена р27, белок р27 ингибирует целый ряд комплексов циклин-ЦЗК и одновременно активирует ген Cdc25a, продукт которого, специфическая фосфатаза, активирует киназы и ЦЗК2 путем их фосфорилирования. Данные процессы приводят к: увеличению содержания в клетке циклина Д и реагирующих с ним киназ ЦЗК4, ЦЗК6; снижению содержания ряда ингибиторов киназ (Cdk); Все это обеспечивает формирование в клетке достаточного количества активных комплексов циклина Д - ЦЗК4,6. Таким образом, комплексы G1 обеспечивают: а) процессы, имеющие место в G1- фазе; б) синтез факторов и ферментов, участвующих в процессе репликации ДНК 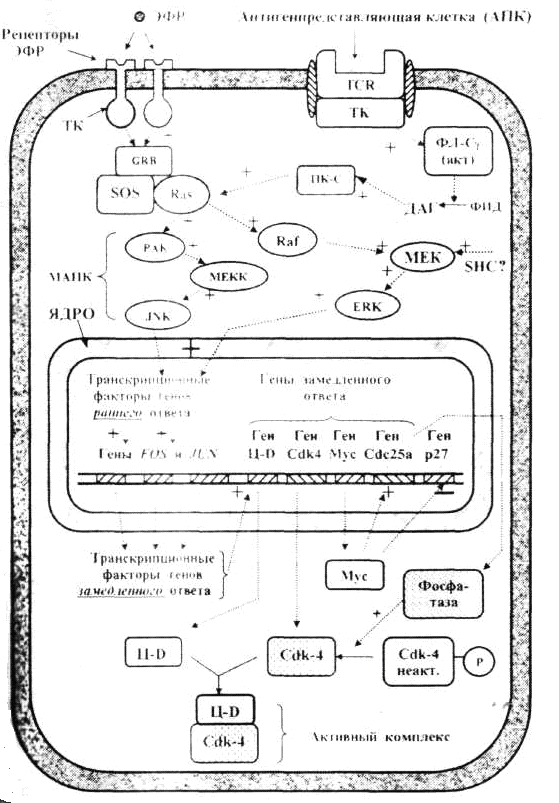 Рис.2. Сигнальные пути, инициирующие пролиферацию: а) стволовых эпителиоцитов (начало – от ЭФР); б) Т-хелперов (начало – от взаимодействия с АПК). Действие комплексов S-и G2-периодов. Комплексами S-периода являются ЦА-Cdk2 и ЦВ-Cdk2. Основная задача этих комплексов – обеспечить прохождение репликации т.е., чтобы каждый участок ДНК был реплицирован только один раз. В S-фазе в цитоплазме происходит удвоение центриолей. В G2-периоде происходит накопление MPF (митоз стимулирующий фактор –mitosis-promoting factor). MPF представляет собой комплекс циклин В-Cdk1. В G2-периоде синтезируется белок тубулин для образования веретена деления. Регуляция митоза На стадиях профазы и метафазы ключевую роль играет высокая активность фактора МРF. Для прохождения анафазы необходимо разрушение MPF. Это осуществляется фактором, обеспечивающим анафазу (АРС –синтезируется в G1 периоде). В метафазу митоза MPF фосфорилирует помимо других белков и APC, который активируется и начинает разрушать молекулы циклина В, входящего в состав MPF, и MPF разрушается в результате фосфорилированные в профазе и метафазе белки дефосфорилируются. Действие этих белков приводит к восстановлению клеточных структур и цитотомии (цитокинезу). Наряду со стимуляторами митоза существуют факторы, препятствующие вступлению клетки в митоз – антимитогенные факторы: 1) фактор некроза опухолей (ФНО) – способствует торможению каскада МАПК, вследствие этого снижается количество активных комплексов циклина Д - ЦЗК4,6. 2) трансформирующий фактор роста (ТФР), запускают механизмы каскадов цитоплазматических белков семейства - smad. Этот комплекс выполняет роль транскрипционного фактора для генов, кодирующих синтез белков р15 и р21, ингибиторов киназ-ЦЗК. Накопление в клетке этих ингибиторов приводит к торможению пролиферации. 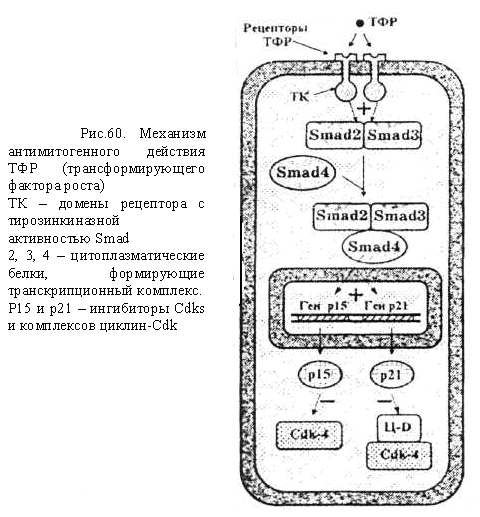 Чекпойнты, контролирующие повреждения ДНК Одновременно с процессами, контролирующими и регулирующими митотический цикл, в клетке имеет место самоконтроль собственного состояния. Контроль за правильностью протекания репликации и распределения генетического материала осуществляется на четырех этапах митотического цикла (G1, S, G2 и М-фазы) и контролируются специальными клеточными механизмами (белком р53). Пункты проверки называются сверочными точками (check-points). Таблица: Характеристика сверочных точек
Если в любой из этих фаз обнаруживается повреждения, клетка задерживается в соответствующих фазах до тех пор, пока поврежденная ДНК не будут восстановлены ферментами репарации. Задержка в фазах предотвращает копирование поврежденных оснований и тормозит мутацию ДНК. Центральную роль в остановке цикла играет белок р53. Он синтезируется в клетке постоянно, но очень быстро разрушатся. При устранении неполадок в клетке концентрация белка р53 снижается, ингибитор Cdk отделяется, и Cdk начинает снова экспрессироваться. Таким образом, в зависимости от результатов «проверки», выбирается один из трех вариантов: а) при отсутствии повреждений – безостановочный переход к следующей стадии цикла; б) задержка на текущей стадии для исправления обнаруженных дефектов; в) запуск механизма апоптоза, если выявленные нарушения неисправимы |