днк. Днк. Затем в 1935 году в Ученых записках мгу
 Скачать 246.81 Kb. Скачать 246.81 Kb.
|

|
Днк ДНК как химическое вещество была выделена Иоганном Фридрихом Мишером в 1869 году из остатков клеток, содержащихся в гное. Он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, вещество получило название нуклеиновая кислота[4]. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Более того, даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию. До 1930-х годов считалось, что ДНК содержится только в животных клетках, а в растительных — РНК. В 1934 году в журнале «Hoppe-Seyler’s Zeitschrift fur physiologishe Chemie»[5], затем в 1935 году в «Ученых записках МГУ»[6] вышли статьи советских биохимиков А. Н. Белозерского и А. Р. Кизеля, в которых доказывалось присутствие ДНК в растительных клетках. В 1936 году группой Белозерского ДНК была выделена из семян и тканей бобовых, злаковых и других растений[7]. Результатом исследований этой же группы советских учёных в 1939 — 1947 годах стала первая в мировой научной литературе информация о содержании нуклеиновых кислот у различных видов бактерий. Постепенно было доказано, что именно ДНК, а не белки, как считалось раньше, является носителем генетической информации. Одно из первых решающих доказательств принесли эксперименты Освальда Эвери, Колина Маклауда и Маклина Маккарти (1944 г.) по трансформации бактерий. Им удалось показать, что за так называемую трансформацию (приобретение болезнетворных свойств безвредной культурой в результате добавления в неё мёртвых болезнетворных бактерий) отвечает выделенная из пневмококков ДНК. Эксперимент американских учёных Алфреда Херши и Марты Чейз (эксперимент Херши — Чейз, 1952 г.) с помеченными радиоактивными изотопами белками и ДНК бактериофагов показали, что в заражённую клетку передаётся только нуклеиновая кислота фага, а новое поколение фага содержит такие же белки и нуклеиновую кислоту, как исходный фаг[8]. Вплоть до 50-х годов XX века точное строение ДНК, как и способ передачи наследственной информации, оставалось неизвестным. Хотя и было доподлинно известно, что ДНК состоит из нескольких цепочек, состоящих из нуклеотидов, никто не знал точно, сколько этих цепочек и как они соединены. В результате работы группы биохимика Эрвина Чаргаффа в 1949—1951 гг. были сформулированы так называемые правила Чаргаффа. Чаргаффу и сотрудникам удалось разделить нуклеотиды ДНК при помощи бумажной хроматографии и определить точные количественные соотношения нуклеотидов разных типов. Соотношение, выявленное для аденина (А), тимина (Т), гуанина (Г) и цитозина (Ц), оказалось следующим: количество аденина равно количеству тимина, а гуанина — цитозину: А=Т, Г=Ц[9][10]. Эти правила, наряду с данными рентгеноструктурного анализа, сыграли решающую роль в расшифровке структуры ДНК. Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин, и правил Чаргаффа[11]. Позже предложенная Уотсоном и Криком модель строения ДНК была доказана, а их работа отмечена Нобелевской премией по физиологии или медицине 1962 г. Среди лауреатов не было скончавшейся к тому времени от рака Розалинд Франклин, так как премия не присуждается посмертно[12]. Интересно, что в 1957 году американцы Александер Рич, Гэри Фелзенфелд и Дэйвид Дэйвис описали нуклеиновую кислоту, составленную тремя спиралями[13]. А в 1985—1986 годах Максим Давидович Франк-Каменецкий в Москве показал, как двухспиральная ДНК складывается в так называемую H-форму, составленную уже не двумя, а тремя нитями ДНК[14][15]. Структура молекулы[править | править код] Нуклеотиды[править | править код]
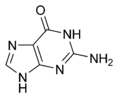
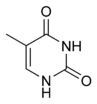
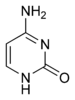
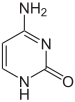
Структуры оснований в составе ДНК Дезоксирибонуклеиновая кислота (ДНК) представляет собой биополимер (полианион), мономером которого является нуклеотид[16][17]. Каждый нуклеотид состоит из остатка фосфорной кислоты, присоединённого по 5'-положению к сахару дезоксирибозе, к которому также через гликозидную связь (C—N) по 1'-положению присоединено одно из четырёх азотистых оснований. Именно наличие характерного сахара и составляет одно из главных различий между ДНК и РНК, зафиксированное в названиях этих нуклеиновых кислот (в состав РНК входит сахар рибоза)[18]. Пример нуклеотида — аденозинмонофосфат, у которого основанием, присоединённым к фосфату и рибозе, является аденин (A) (показан на рисунке). Исходя из структуры молекул, основания, входящие в состав нуклеотидов, разделяют на две группы: пурины (аденин [A] и гуанин [G]) образованы соединёнными пяти- и шестичленным гетероциклами; пиримидины (цитозин [C] и тимин [T]) — шестичленным гетероциклом[19]. В виде исключения, например, у бактериофага PBS1, в ДНК встречается пятый тип оснований — урацил ([U]), пиримидиновое основание, отличающееся от тимина отсутствием метильной группы на кольце, обычно заменяющее тимин в РНК[20]. Тимин (T) и урацил (U) не так строго приурочены к ДНК и РНК соответственно, как это считалось ранее. Так, после синтеза некоторых молекул РНК значительное число урацилов в этих молекулах метилируется с помощью специальных ферментов, превращаясь в тимин. Это происходит в транспортных и рибосомальных РНК[21]. Двойная спираль[править | править код]  В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует в разных формах. На рисунке представлены формы A, B и Z (слева направо) Полимер ДНК обладает довольно сложной структурой. Нуклеотиды соединены между собой ковалентно в длинные полинуклеотидные цепи. Эти цепи в подавляющем большинстве случаев (кроме некоторых вирусов, обладающих одноцепочечными ДНК-геномами) попарно объединяются при помощи водородных связей во вторичную структуру, получившую название двойной спирали[11][18]. Остов каждой из цепей состоит из чередующихся фосфатов и сахаров[22]. Внутри одной цепи ДНК соседние нуклеотиды соединены фосфодиэфирными связями, которые формируются в результате взаимодействия между 3'-гидроксильной (3'—ОН) группой молекулы дезоксирибозы одного нуклеотида и 5'-фосфатной группой (5'—РО3) другого. Асимметричные концы цепи ДНК называются 3' (три прайм) и 5' (пять прайм). Полярность цепи играет важную роль при синтезе ДНК (удлинение цепи возможно только путём присоединения новых нуклеотидов к свободному 3'-концу). Как уже было сказано выше, у подавляющего большинства живых организмов ДНК состоит не из одной, а из двух полинуклеотидных цепей. Эти две длинные цепи закручены одна вокруг другой в виде двойной спирали, стабилизированной водородными связями, образующимися между обращёнными друг к другу азотистыми основаниями входящих в неё цепей. В природе эта спираль, чаще всего, правозакрученная. Направления от 3'-конца к 5'-концу в двух цепях, из которых состоит молекула ДНК, противоположны (цепи «антипараллельны» друг другу). Ширина двойной спирали составляет от 22 до 24 Å, или 2,2—2,4 нм, длина каждого нуклеотида 3,3 Å (0,33 нм)[23]. Подобно тому, как в винтовой лестнице сбоку можно увидеть ступеньки, на двойной спирали ДНК в промежутках между фосфатным остовом молекулы можно видеть рёбра оснований, кольца которых расположены в плоскости, перпендикулярной по отношению к продольной оси макромолекулы. В двойной спирали различают малую (12 Å) и большую (22 Å) бороздки[24]. Белки, например, факторы транскрипции, которые присоединяются к определённым последовательностям в двухцепочечной ДНК, обычно взаимодействуют с краями оснований в большой бороздке, где те более доступны[25]. Образование связей между основаниями[править | править код] См. также: Полимеразная цепная реакция Каждое основание на одной из цепей связывается с одним определённым основанием на второй цепи. Такое специфическое связывание называется комплементарным. Пурины комплементарны пиримидинам (то есть способны к образованию водородных связей с ними): аденин образует связи только с тимином, а цитозин — с гуанином. В двойной спирали цепочки также связаны с помощью гидрофобных взаимодействий и стэкинга, которые не зависят от последовательности оснований ДНК[26]. Комплементарность двойной спирали означает, что информация, содержащаяся в одной цепи, содержится и в другой цепи. Обратимость и специфичность взаимодействий между комплементарными парами оснований важна для репликации ДНК и всех остальных функций ДНК в живых организмах. Так как водородные связи нековалентны, они легко разрываются и восстанавливаются. Цепочки двойной спирали могут расходиться как замок-молния под действием ферментов (хеликазы) или при высокой температуре[27]. Разные пары оснований образуют разное количество водородных связей. АТ связаны двумя, ГЦ — тремя водородными связями, поэтому на разрыв ГЦ требуется больше энергии. Процент ГЦ-пар и длина молекулы ДНК определяют количество энергии, необходимой для диссоциации цепей: длинные молекулы ДНК с большим содержанием ГЦ более тугоплавки[28]. Температура плавления нуклеиновых кислот зависит от ионного окружения, рост ионной силы стабилизирует ДНК по отношению к денатурированию. При добавлении к ДНК хлорида натрия существует линейная зависимость между температурой плавления и логарифмом ионной силы раствора. Предполагается, что добавление электролита ведет к экранированию зарядов в цепях ДНК и этим уменьшает силы электростатического отталкивания между заряженными фосфатными группами, способствуя жесткости структуры. Аналогично температуру плавления ДНК повышают ионы марганца, кобальта, цинка и никеля, но ионы меди, кадмия и свинца, напротив, понижают ее[29]. Части молекул ДНК, которые из-за их функций должны быть легко разделяемы, например, ТАТА последовательность в бактериальных промоторах, обычно содержат большое количество А и Т. Химические модификации азотистых оснований[править | править код] Основная статья: Метилирование ДНК
Структура цитозина, 5-метилцитозина и тимина. Тимин может возникать путём деаминирования 5-метилцитозина Азотистые основания в составе ДНК могут быть ковалентно модифицированы, что используется при регуляции экспрессии генов. Например, в клетках позвоночных метилирование цитозина с образованием 5-метилцитозина используется соматическими клетками для передачи профиля генной экспрессии дочерним клеткам. Метилирование цитозина не влияет на спаривание оснований в двойной спирали ДНК. У позвоночных метилирование ДНК в соматических клетках ограничивается метилированием цитозина в последовательности ЦГ[30]. Средний уровень метилирования отличается у разных организмов, так, у нематоды Caenorhabditis elegans метилирование цитозина не наблюдается, а у позвоночных обнаружен высокий уровень метилирования — до 1 %[31]. Другие модификации оснований включают метилирование аденина у бактерий и гликозилирование урацила с образованием «J-основания» в кинетопластах[32]. Метилирование цитозина с образованием 5-метилцитозина в промоторной части гена коррелирует с его неактивным состоянием[33]. Метилирование цитозина важно также для инактивации Х-хромосомы у млекопитающих[34]. Метилирование ДНК используется в геномном импринтинге[35]. Значительные нарушения профиля метилирования ДНК происходят при канцерогенезе[36]. Несмотря на биологическую роль, 5-метилцитозин может спонтанно утрачивать аминную группу (деаминироваться), превращаясь в тимин, поэтому метилированные цитозины являются источником повышенного числа мутаций[37]. Повреждения ДНК[править | править код] Основная статья: Мутация  Интеркалированное химическое соединение, которое находится в середине спирали — бензопирен, основной мутаген табачного дыма[38] ДНК может повреждаться разнообразными мутагенами, к которым относятся окисляющие и алкилирующие вещества, а также высокоэнергетическая электромагнитная радиация — ультрафиолетовое и рентгеновское излучение. Тип повреждения ДНК зависит от типа мутагена. Например, ультрафиолет повреждает ДНК путём образования в ней димеров тимина, которые возникают при образовании ковалентных связей между соседними основаниями[39]. Оксиданты, такие как свободные радикалы или пероксид водорода, приводят к нескольким типам повреждения ДНК, включая модификации оснований, в особенности гуанозина, а также двухцепочечные разрывы в ДНК[40]. По некоторым оценкам, в каждой клетке человека окисляющими соединениями ежедневно повреждается порядка 500 оснований[41][42]. Среди разных типов повреждений наиболее опасные — это двухцепочечные разрывы, потому что они трудно репарируются и могут привести к потерям участков хромосом (делециям) и транслокациям. Многие молекулы мутагенов вставляются (интеркалируют) между двумя соседними парами оснований. Большинство этих соединений, например: бромистый этидий, даунорубицин, доксорубицин и талидомид, имеет ароматическую структуру. Для того чтобы интеркалирующее соединение могло поместиться между основаниями, они должны разойтись, расплетая и нарушая структуру двойной спирали. Эти изменения в структуре ДНК мешают репликации, вызывая мутации, и транскрипции. Поэтому интеркалирующие соединения часто являются канцерогенами, наиболее известные из которых — бензопирен, акридины, афлатоксин и бромистый этидий[43][44][45]. Несмотря на эти негативные свойства, в силу их способности подавлять транскрипцию и репликацию ДНК, интеркалирующие соединения используются в химиотерапии для подавления быстро растущих клеток рака[46]. Некоторые вещества (цисплатин[47], митомицин C[48], псорален[49]) образуют поперечные сшивки между нитями ДНК и подавляют синтез ДНК, благодаря чему используются в химиотерапии некоторых видов рака ( |