|
|
Колок. Знать эволюционные связи (филогенез органов, возникновение пороков развития)
Мембранный транспорт- транспорт веществ сквозь клеточную мембрану в клетку или из клетки, осуществляемый с помощью различных механизмов - простой диффузии, облегченной диффузии и активного транспорта.
Важнейшее свойство биологической мембраны состоит в ее способности пропускать в клетку и из нее различные вещества.Это имеет большое значение для саморегуляции и поддержания постоянного состава клетки. Такая функция клеточной мембраны выполняется благодаря избирательной проницаемости, т.е. способностью пропускать одни вещества и не пропускать другие.
11.
Клеточное ядро состоит из оболочки, ядерного сока, ядрышка и хроматина. Функциональная роль ядерной оболочки заключается в обособлении генетического материала (хромосом) эукариотической клетки от цитоплазмы с присущими ей многочисленными метаболическими реакциями, а также регуляции двусторонних взаимодействий ядра и цитоплазмы. Ядерная оболочка состоит из двух мембран, разделенных околоядерным (перинуклеарным) пространством. Последнее может сообщаться с канальцами цитоплазматической сети. Ядерная оболочка пронизана порожу диаметром 80—90 нм. Область поры или поровый комплекс с диаметром около 120 нм имеет определенное строение, что указывает на сложный механизм регуляции ядерно-цитоплазматических перемещений веществ и структур. Количество пор зависит от функционального состояния клетки. Чем выше синтетическая активность в клетке, тем больше их число. Основу ядерного сока, или матрикса, составляют белки. Ядерный сок образует внутреннюю среду ядра, в связи с чем он играет важную роль в обеспечении нормального функционирования генетического материала. Ядрышко представляет собой структуру, в которой происходит образование и созревание рибосомальных РНК (рРНК). Гены рРНК занимают определенные участки (в зависимости от вида животного) одной или нескольких хромосом (у человека 13—15 и 21—22 пары) — ядрышковые организаторы, в области которых и образуются ядрышки.
12.
В цитоплазме различают основное вещество (матрикс, гиалоплазма), включения и органеллы. Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. Органеллы — это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции.
Выделяют органеллы общего значения и специальные. Последние в значительном количестве присутствуют в клетках, специализированных к выполнению определенной функции, но в незначительном количестве могут встречаться и в других типах клеток. К ним относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки. К органеллам общего значения относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. В растительных клетках выделяют также хлоропласта, в которых происходит фотосинтез.
13.
а) болезни, вызванные точковыми мутациями, приводящими к замене консервативных аминокислот в собственных белках митохондрий. К ним относятся пигментный ретинит и нейроофтальмопатия Лебера, при которой наступает двусторонняя потеря зрения. Выраженность клинических признаков у больных этими заболеваниями коррелирует с количеством мутантной мтДНК, которое у разных больных может варьировать от 5 до 100% всей мтДНК;
б) болезни, вызванные мутациями в генах т-РНК, приводящими к многочисленным дегенеративным заболеваниям с различной степенью тяжести клинических проявлений, коррелирующей с количеством мутантной мтДНК;
в) болезни вызванные делениями и дупликациями участков митохондриалъных генов. У человека описано тяжелое заболевание молодого и среднего возраста — отсроченная кардиопатия, при которой обнаружены делеции мтДНК кардиоцитов. Заболевание носит семейный характер. В ряде случаев предполагается Х-сцепленное наследование, что позволяет думать о существовании ядерного гена, мутация которого вызывает делению до 50% мтДНК кардиоцитов;
+г) болезни, вызванные снижением числа копий мтДНК, что является следствием определенных мутаций. К данной группе относятся летальная инфантильная дыхательная недостаточность и синдром молочнокислого ацидоза, при которых число копий мтДНК снижается до 1—2% от нормы. Снижение содержания мтДНК в клетках различных органов приводит к развитию миопатий, нефропатий, печеночной недостаточности и т.д. вследствие ослабления синтеза белков, кодируемых мтДНК.
Пероксисомные болезни — группа заболеваний, обусловленных нарушением структуры и функции пероксисом, - внутриклеточных органелл, присутствующих в каждой клетке организма за исключением зрелых эритроцитов. Одна из важнейших функций пероксисом — зашита клетки от образующегося в ней атомарного кислорода с помощью химических превращений, в которых участвуют пероксисомные каталазы. Клиническая картина пероксисомных заболеванийвключает: 1) черспно-лицевые аномалии; 2) патологию органов зрения (нарушение пигментации сетчатки, побледнение дисков зрительных нервов); 3) неврологические симптомы (мышечная гипотония, судороги, задержка раннего психомоторного развития); 4) гепатомегалию; 5) аномалии скелета (ризомелический тип укорочения конечностей).
14. Панспермия - это астробиологическая гипотеза, согласно которой жизнь существует где-то еще во Вселенной из-за распространения микроорганизмов, которые случайно переносятся на астероидах, других путешествующих планетоидах или даже на космических кораблях. Гипотезы панспермии могут позволить нам объяснить абиогенез или, другими словами, происхождение жизни и то, как она может перемещаться из мест, где обитают живые организмы, чтобы заселить далекие галактики или планеты.
Во-первых, это механизм радиопанспермии, при котором микроорганизмы перемещаются в космосе благодаря радиационному давлению звезд, пока не достигнут объекта, на котором условия могут быть благоприятными или нет, чтобы он мог развиться в более крупные и сложные формы жизни. Однако эта гипотеза была опровергнута из-за длительного воздействия радиации на бактерии во время межпланетных путешествий, которая денатурирует любую форму ДНК или РНК и снижает любые шансы на успешную радиопанспермию почти до нуля. Вторая - гипотеза литопансмермии. В этом случае межпланетный перенос жизни будет происходить благодаря присутствию микроорганизмов на астероидах, кометах и других планетоидах или в них, достаточно больших и устойчивых, чтобы выдержать планетарный выброс, длительные межгалактические, планетные или звездные путешествия, а также атмосферные повторный вход. Наконец, гипотезы о направленной панспермии, или намеренной реализации жизни на Земле, также считаются правдоподобными. По мнению лауреата Нобелевской премии Фрэнсиса Крика, жизнь на Земле могла возникнуть в результате "межзвездного осеменения" нашей планеты развитой внеземной цивилизации.
Первым условием выживания организмов в открытом космосе является их способность выживать в отсутствие солнечного света. Однако вплоть до 1970-х годов такие живые организмы были неизведанными, пока в глубинах Галапагосского разлома не были обнаружены организмы, для существования которых требовались только вода и энергия, синтезированные из окисления реактивных химических веществ, обнаруженных в почве Земли. Это пролило свет на новую категорию бактерий, ныне известных как экстремофилы. У этих микроорганизмов развиваются цисты и споры, чтобы защитить себя при воздействии сильного стресса (гамма-излучение, температура, ультрафиолет…). Однако после восстановления жизнеспособных условий эти споры и цисты прорастут и позволят организму снова стать метаболически активным.
Гипотеза биохимической эволюции Опарина–Холдейна (гипотеза абиогенеза)
Возникновение жизни на нашей планете произошло в несколько этапов эволюции:
Абиогенный синтез простых органических соединений.
Образование биополимеров.
Установление связей между биополимерами — образование коацерватов.
Возникновение мембран, отделяющих первые подобия живых организмов — протобионтов — от окружающей среды.
Возникновение обмена веществ и энергии с окружающей средой.
Появление способности к самовоспроизведению.
Формирование экологических связей и образование первых экосистем.
15. ДНК — макромолекула, обеспечивающая хранение, передачу из поколения в поколение и
Реализацию генетической программы развития и функционирования живых организмов.
Основная роль ДНК в клетках — долговременное хранение информации о структуре РНК и белков.
Важнейшее свойство ДНК —комплементарность ее цепей. Это означает, что против аденина в одной из цепей всегда стоит Тимин в другой цепи, гуанин всегда соединен с цитозином. Комплементарные пары аденин и тимин соединены двумя водородными связями, а гуанин с цитозином тремя водородными связями.
Помимо водородных связей между основаниями разных цепей стабильность двойной
спирали ДНК обеспечивают гликозидные связи между азотистыми основаниями и остатками
дезоксирибозы, а также фосфодиэфирные связи между двумя соседними остатками
дезоксирибозы.
Особенности ДНК:
1. Стабильность. Она обеспечивается водородными, гликозидными и фосфодиэфирными
связями, а также механизмом репарации спонтанных и индуцированных повреждений;
2. Способность к репликации. Благодаря этому механизму в соматических клетках
сохраняется диплоидное число хромосом. Схематично псе перечисленные особенности ДНК
как генетической молекулы изображены на рисунке.
3. Наличие генетического кода. Последовательность оснований в ДНК с помощью
процессов транскрипции и трансляции преобразуется в последовательность аминокислот в
полипептидной цепи;
4. Способность к генетической рекомбинации. Благодаря этому механизму образуются
новые сочетания сцепленных генов.
Передача генетической информации в клетке основана на матричных процессах (репликации, транскрипции, трансляции). Синтез дочерней цепи (репликация) молекулы ДНК происходит по матрице одной из двух родительских цепей с образованием новой двуцепочечной молекулы ДНК. Синтез молекулы РНК совершается в процессе транскрипции ДНК по матрице одной из двух цепей ДНК. Такая матричная(информационная) РНК может рассматриваться как посредник между ДНК и белком. Далее при синтезе белков генетическая информация, закодированная в последовательности триплетов азотистых оснований (канонов), транслируется в аминокислотную последовательность полипептидных цепей.
Репликация ДНК – это молекулярный процесс точного копирования молекул ДНК (ее нуклеотидной последовательности). С помощью механизма репликации происходит точная передача генетической информации от клетки к клетке и, таким образом, все клетки многоклеточного организма являются носителями одной и той же наследственной информации. Процесс синтеза ДНК сопровождается множеством событий и является, как правило, точным. Из одной молекулы ДНК синтезируются две идентичные дочерние молекулы. Этот процесс становится возможным благодаря структурным особенностям молекулы ДНК: - двуцепочечная структура; - комплементарность и антипараллельность. Для начала репликации ДНК фрагментов этой нити требуется синтез коротких фрагментов РНК (о характерных особенностях РНК будет сказано ниже) как затравок, для чего используется особый фермент — РНК-полимераза (праймаза). Впоследствии праймеры РНК удаляются, в образовавшиеся бреши встраивается ДНК с помощью ДНК полимеразы I. Таким образом, каждая цепь ДНК используется как матрица или шаблон для построения комплементарной цепи, и репликация ДНК является полуконсервативной (т.е. одна нить в новой молекуле ДНК — «старая», а вторая — новая).
Для репликации, лидирующей и отстающей цепей клеткой используют разные ферменты. В результате репликации образуются две новые абсолютно идентичные молекулы ДНК, идентичные также исходной молекуле ДНК до начала ее редупликации (более подробно процесс репликации ДНК показан на рис. 3.5). ДНК-полимераза, как и любой другой фермент, существенно ускоряет процесс присоединения комплементарных нуклеотидов к свободной цепи ДНК, однако химическое сродство аденина к тимину, а цитозина к гуанину столь велико, что они соединяются друг с другом и в отсутствие ДНК-полимеразы в простой реакционной смеси. Полуконсервативный механизм репликации предполагает разделение цепей исходной молекулы ДНК. При этом каждая материнская цепь является матрицей для синтеза дочерней цепи. Таким образом, при репликации образуются две молекулы ДНК, каждая из которых включает одну материнскую цепь и одну новую — дочернюю.
16.
В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1. Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2. Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3. Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (
10 пн) некодирующие последовательности, которые входят в состав прицентромерного гетерохроматина.
17.
. Хроматин (греч. chroma — цвет, краска и греч. nitos — нить) — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоида у прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК. Основную массу хроматина составляют белки гистоны. Гистоны являются компонентом нуклеосом, — надмолекулярных структур, участвующих в упаковке хромосом.
эухроматин, локализующийся ближе к центру ядра, более светлый, более деспирилизованный, менее компакт-ный, более активен в функциональном отношении. Эухроматин - неконденсированный хроматин, с которого происходит синтез белка.
Гетерохроматин - конденсированный хроматин, с которого белок не синтезируется. гетерохроматин - плотно спирализованная часть хроматина. Гетерохроматин соответствует конденсированным, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипции). Он интенсивно окра-шивается основными красителями, и в световом микроскопе имеет вид тёмных пятен, гранул.
18. Хромосомы — это структуры ядра, которые представляют собой максимально спирализованный наследственный материал клетки. Хромосомы исследуют в прометафазу или метафазу (метафазные пластинки) митоза
Функции:
1) хранение наследственной информации2)
использование наследственной информации для создания и поддержания клеточной организации.
3) регуляция считывания наследств информации
4)размножение н.и. путем редупликации
5)передача н.и.
Химический состав:
1)ДНК 2) гистоновые белки 3) негистоновые (кислые) белки 4) РНК 5) ионы металлов 6) липиды, полисахариды.
Метафазные хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. На стадии метафазы сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначается буквой p, длинное плечо — буквой q.
Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом. Морфология хромосом Хромосомы могут находиться в двух структурно-функциональных состояниях : в конденсированном (спирализованном) и деконденсированном (деспирализованном). Чем более диффузен (деспирализован) хроматин, тем интенсивнее синтетичесике процессы в клетке. И наоборот, чем более спирализованы хромосомы, тем ближе процесс деления клетки. Морфологически в хромосоме различают: 1) первичную перетяжку – центромеру (кинетохор), к кот. прикрепляются нити веретена деления 2) плечи хромосом 3) вторичную перетяжку (ядрышковый организатор) 4) спутник (сателлит) По расположению первичной перетяжки хромосомы делятся на: метацентрические (равноплечие), субметацентрические ( неравноплечие), акроцентрические – одно плечо гораздо короче другого и телоцентрические (одно плечо отсутствует).
19.
Период существования клетки от момента ее образования путем деления материнской клетки (включая само деление) до собственного деления или смерти называютжизненным (клеточным) циклом.Продолжительность жизненного цикла различных клеток многоклеточного организма различна. Так, клетки нервной ткани после завершения эмбрионального периода перестают делиться и функционируют н а протяжении всей жизни организма, а затем погибают. Митоз –непрямое деление соматических клеток, в результате которого сначала происходит удвоение, а затем равномерное распределение наследственного материала между дочерними клетками. Биологическое значение митоза: в результате митоза образуются две клетки, каждая из которых содержит столько же хромосом, сколько их было в материнской. Митотический циклсостоит из интерфазы и митоза. Длительность митотического цикла у разных организмов сильно варьирует. Непосредственно на деление клетки уходит обычно 1–3 ч, то есть основную часть жизни клетка находится в интерфазе. Начальный отрезок интерфазы – пресинтетический период(2n2с, где n – количество хромосом, с – количество ДНК),период роста,начинающийся непосредственно после митоза.Синтетический периодпо продолжительности очень различен: от нескольких минут у бактерий до 6–12 ч в клетках млекопитающих. Во время синтетического периода происходит самое главное событие интерфазы – удвоение молекул ДНК. Каждая хромосома становится двухроматидной, а число хромосом не изменяется (2n4с).
Постсинтетический период.Обеспечивает подготовку клетки к делению и также характеризуется интенсивными процессами синтеза белков, входящих в состав хромосом; синтезируются ферменты и энергетические вещества, необходимые для обеспечения процесса деления клетки.
20. Репарация — особая функция клеток, заключающаяся в способности исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК в клетке или в результате воздействия физических или химических агентов. Осуществляется специальными ферментными системами клетки. Виды репорации:
1. Эксцизионная репарация восстанавливает поврежденные участки как УФ-облучением, так и другими факторами. Эта система репарации имеет несколько ферментов: репарационные эндонуклеаза и экзонуклеаза, ДНК-полимераза, ДНК-лигаза.
2. Пострепликативная репарация является неполной, так как идет «в обход», и поврежденный участок из молекулы ДНК не удаляется.
3. Прямая репарация — наиболее простой путь устранения повреждений в ДНК, в котором обычно задействованы специфические ферменты, способные быстро устранять соответствующее повреждение, восстанавливая исходную структуру нуклеотидов.
21. Регуляция клеточной активности. Гибель клеток – как нормальный физиологический процесс.
Гибель клетки — необратимый процесс. Он развивается вследствие её значительного повреждения (некроз) или активации специализированной программы смерти (апоптоз).
Некроз — гибель клетки в результате воздействия на неё повреждающего фактора. Этот фактор приводит к изменениям клетки, исключающим её дальнейшее существование. Понятие «некроз» рассматривается как синоним термина «гибель клетки от повреждения». Одним из основных механизмов гибели клетки является лизис. Этот термин обычно применяют в случаях гибели клетки вследствие первичного и массированного повреждения её плазматической мембраны, что приводит к смертельному для клетки, практически мгновенному нарушению её ионного состава (например, лизис инфицированной вирусом клетки; эритроцитов в гипотонической среде; раковой клетки, атакованной цитотоксическими Т-лимфоцитами).
Апоптоз— гибель клетки вследствие реализации программы, приводящей к поэтапному прекращению её жизнедеятельности (программа смерти клетки предсуществует; её запускает сигнал, чаще внеклеточный). В отличие от некроза, апоптоз — не обязательно патологический процесс. Существует много физиологических процессов (наблюдающихся в основном в ходе морфогенеза или поддержания нормального состава обновляющихся клеточных популяций), когда клетки, завершившие свой жизненный цикл, удаляются путём апоптоза.
22. Размножение - это способность всех организмов воспроизводить себе подобных, что обеспечивает непрерывность и приемлемость жизни. 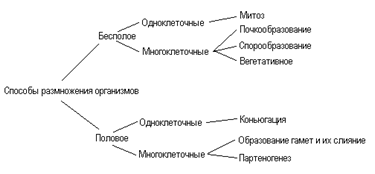
23. Бесполое размножение – лежит деление клеток путем митоза, при котором
из каждой материнской клетки создаются две равноценные дочерние
клетки (два организма).
Биологическая роль бесполого размножения:
1. Возникновении организмов, идентичных родительским по содержанию наследственного материала, а также анатомическим и физиологическим свойствам (биологические копии).
Способы бесполого размножения:
1. Деление — способ бесполого размножения, характерный для одноклеточных организмов, при котором материнская особь делится на две или большее количество дочерних клеток.
2. Почкование — способ бесполого размножения, при котором новые особи образуются в виде выростов на теле родительской особи. Дочерние особи могут отделяться от материнской и переходить к самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом случае колонии (коралловые полипы).
3. Фрагментация — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды, спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации.
4. Полиэмбриония — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается эмбрион (монозиготные близнецы).
5. Вегетативное размножение — способ бесполого размножения, при котором новые особи образуются или из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.), специально предназначенных для этой формы размножения. Вегетативное размножение характерно для многих групп растений, используется в садоводстве, огородничестве, селекции растений (искусственное вегетативное размножение).
6. Спорообразование — размножение посредством спор. Споры— специализированы клетки, у большинства видов образуются в особых органах — спорангиях. У высших растений образованию спор предшествует мейоз.
7. Клонирование— комплекс методов, используемых человеком для получения генетически идентичных копий клеток или особей. Клон— совокупность клеток или особей, произошедших от общего предка путем бесполого размножения. В основе получения клона лежит митоз (у бактерий — простое деление).
Половое размножение — это способ размножения, при котором новый организм формируется в результате комбинации генов, принадлежащих обоим родителям.
Формы полового размножения:
1. Оплодотворение — это процесс слияния половых клеток родительских организмов с образованием зиготы. Таким образом, мейоз, уменьшающий набор хромосом, и оплодотворение, восстанавливающее его, являются основой видового постоянства числа хромосом. Многие организмы являются разнополыми, что обеспечивает новую комбинацию генов родителей у потомков. Однако даже гермафродитные организмы, продуцирующие оба типа половых клеток, имеют специальные механизмы, препятствующие самооплодотворению.
2. Конъюгация — это слияние наследственной информации двух неспециализированных клеток или обмен ею. Она характерна для инфузорий, некоторых водорослей и грибов.
3. Партеногенез - процесс, при котором не требуется оплодотворения, а новый организм формируется из неоплодотворенной яйцеклетки. Он встречается, например, у рачков-дафний, пчел, некоторых ящериц, ряда растений.
Значение полового размножения:
Оно обеспечивает появление пусть и небольшого числа потомков, но они имеют новые комбинации генов и признаков родителей, что позволяет им приспосабливаться к изменяющимся условиям окружающей среды.
24. Гаметогенез – это последовательный процесс, который обеспечивает размножение, рост и созревание половых клеток в мужском организме (сперматогенез) и женском (овогенез).
Гаметогенез протекает в половых железах - сперматогенез в семенниках у мужчин, а овогенез в яичниках у женщин. В результате гаметогенеза в организме женщины образуются женские половые клетки - яйцеклетки, а у мужчин - мужские половые клетки сперматозоиды.
Именно процесс гаметогенез (сперматогенез, овогенез) дает возможность мужчине и женщине возможность воспроизведения потомства.
Мейоз — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза.
Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
Профаза I — профаза первого деления очень сложная и состоит из 5 стадий:
1. Лептотена или лептонема — упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются).
2. Зиготена или зигонема — происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация.
3. Пахитена или пахинема — (самая длительная стадия) — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер — обмен участками между гомологичными хромосомами.
4. Диплотена или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток.
5. Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
К концу Профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления, разрушаются ядерная мембрана и ядрышки.
Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки.
Анафаза I — микротрубочки сокращаются, биваленты делятся и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе.
Телофаза I — хромосомы деспирализуются и появляется ядерная оболочка.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления, перпендикулярное первому веретену.
Метафаза II — унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку.
Анафаза II — униваленты делятся и хроматиды расходятся к полюсам.
Телофаза II — хромосомы деспирализуются и появляется ядерная оболочка.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и три так называемых редукционных тельца (абортивные дериваты первого и второго делений).
25. Сперматогенезом называют процесс формирования мужских гамет (половых клеток) - сперматозоидов. Он начинается в период полового созревания (под влиянием мужских половых гормонов) и длится практически до конца жизни. Сперматогенез складывается из четырех фаз (периодов):
Фаза размножения
В ходе фазы размножения диплоидные сперматогенные клетки (2n2c) многократно делятся митозом, в результате образуются сперматогонии (2n2c) - стволовые клетки. Часть сперматогоний вступает в последующее митотическое деление, образуя такие же сперматогонии (2n2c).
Фаза роста
Половые клетки в этой фазе называются сперматоцитами I порядка, они теряют способность к митотическому делению.
В этот период клетка растет, увеличивается количество органоидов и цитоплазмы. Происходит подготовка к мейозу, который начинается в следующей фазе - созревания.
На фазу роста приходится S-период: происходит удвоение ДНК, в результате чего набор хромосом сперматоцита I порядка становится (2n4c).
Фаза созревания
Происходит первое деление мейоза (мейоз I). В результате из сперматоцитов I порядка (2n4c) образуются сперматоциты II порядка (n2c). Между мейозом I и мейозом II практически отсутствует интерфаза, поэтому сперматоциты II порядка (n2c) сразу же вступают в мейоз II, в результате которого образуются сперматиды (nc).
Итак, в фазу созревания происходят первое и второе деления мейоза, которые приводят к тому, что образовавшаяся клетка - сперматида - имеет гаплоидный набор хромосом (nc).
Формирования
В этой фазе у каждой сперматиды отрастает жгутик, после чего они получают полное право называться сперматозоидами. У основания жгутика концентрируются митохондрии - "энергетические станции клетки", которые всегда будут готовы предоставить АТФ для его активной работы.
Оогенезом называют процесс формирования женских гамет (половых клеток) - яйцеклеток. Он активируется в женском организме в период полового созревания (под действием женских половых гормонов) и длится до менопаузы (45-55 лет).
Оогенез протекает по очень похожей со сперматогенезом схеме, однако вы увидите некоторые отличия. Например, фаза формирования, характерная для сперматогенеза, здесь отсутствует, поэтому овогенез складывается из трех фаз:
Фаза размножения
В результате многократных делений клеток яичника образуются стволовые клетки - овогонии (2n2c).
Фаза роста
Половые клетки в этой фазе называются ооцитами I порядка, они теряют способность к митотическому делению.
В овогенезе эта фаза отличается более длительной продолжительностью, по сравнению с такой же фазой в сперматогенезе. Клетки накапливают большой запас питательных веществ. В этот период происходит удвоение ДНК в S-периоде - набор хромосом и ДНК ооцитов I порядка становится 2n4c.
Фаза созревания
Ооциты I порядка (2n4c) вступают в первое деление мейоза, в результате которого образуются ооциты II порядка (n2c) и первое полярное (направительное) тельце, которое не несет большой функциональной значимости и подвергается дегенерации.
Второе деление мейоза начинается только после взаимодействия овоцита II порядка (n2c) со сперматозоидом. В результате этого образуется яйцеклетка (nc) и второе полярное тельце, которое также подвергается дегенерации.
Строго говоря, при овуляции из яичников выходит не "яйцеклетка", а ооцит II порядка, который ждет встречи со сперматозоидом для продолжения деления и развития будущего зародыша. Если такого взаимодействия не происходит, то яйцеклетка подвергается дегенерации.
26. Яйцеклетки – неподвижные, округлой формы клетки. Яйцеклетка — женские гаметы животных, высших растений, а также многих водорослей и других протистов, которым свойственна оогамия. Как правило, яйцеклетки — гаплоидные клетки, но могут иметь другую плоидность у полиплоидных организмов. Человеческая яйцеклетка имеет диаметр примерно 150 мкм. Наиболее очевидная отличительная черта яйцеклетки - это ее большие размеры. Типичная яйцеклетка имеет сферическую или овальную форму, а диаметр ее составляет у человека около 100 мкм (величина типичной соматической клетки около 20 мкм). Столь же внушительными могут быть размеры ядра, в преддверии быстрых делений, следующих сразу за оплодотворением, в ядре откладываются запасы белков. Одной из главных особенностей строения яйцеклеток является формирование у них в процессе овогенеза вторичных, а у птиц и третичных оболочек. Плазмолемма яйцеклеток представляет собой их первичную оболочку, называемую оволеммой. На стадии роста за счет вспомогательных фолликулярных клеток яичников и собственно ооцита образуются вторичные оболочки: неклеточная блестящая (прозрачная) и клеточная – лучистый венец. Ближайшая к оволемме прозрачная оболочка состоит из гликозаминогликанов и белков. Она выполняет защитную функцию, обеспечивает доставку к ооциту питательных веществ, необходимых для осуществления процессов ее жизнедеятельности и формирования желтка; играет важную роль в иммунных взаимодействиях со сперматозоидами и защите яйца от возможного попадания чужеродных антигенов из крови, омывающей фолликулы. Для активизации обменных процессов ооцит и фолликулярные клетки лучистого венца выпячивают в блестящую оболочку, навстречу друг к другу, микроворсинки. Лучистый венец формируется у растущих (вторичных) фолликулов за счет размножения и роста поддерживающих фолликулярных клеток, входящих в состав примордиальных фолликулов. Вследствие гормональной активизации первичные, функционально неактивные вспомогательные клетки делятся митотически, увеличиваются в размерах, меняя свою форму от плоской до кубической и призматической. Высокие призматические клетки, плотно одевая ооцит, формируют вокруг него своеобразную корону из радиально расположенных и самих тел клеток, и их удлиненных ядер. У яиц птиц вторичные оболочки выражены слабо, но зато у них получают особенно сильное развитие третичные оболочки, отсутствующие у млекопитающих. Третичные оболочки формируются за счет активной функции желез разных отделов яйцевода, наслаивающих последовательно свои секреты на уже оплодотворенную яйцеклетку при ее медленном перемещении в клоаку. В результате образуются белковая, наружная и внутренняя подскорлуповые, скорлуповая и надскорлуповая оболочки, выполняющие защитные и трофические функции при развитии
птичьих эмбрионов в достаточно экстремальных для них условиях суши.
Виды:
1. Изолецитальные - микроскопических размеров, содержащая малое количество желтка, который почти равномерно распределен по цитоплазме, ядро располагается примерно в центре; свойственна иглокожим, ланцетнику, живородящим млекопитающим.
2. Телолецитальные -желтка много, и он распределён в цитоплазме яйцеклетки неравномерно. Основная масса желтка скапливается у одного из полюсов-вегетативного; противоположный полюс, к которому оттесняется активная цитоплазма называется анимальным. Если желток всё же погружен в цитоплазму и не обособлен от неё в виде отдельной фракции, как у осетровых и земноводных, яйцеклетки называют умеренно телолецитальными. Если желток полностью отделён от цитоплазмы, как у амниот, то это резко телолецитальные.
3. Центролецитальные- желток расположен в центральной части, в связи с чем Я. ц. претерпевает поверхностное дробление; свойственна насекомым и некоторым Другим членистоногим.
Сперматозоид - мужская половая клетка животных организмов и многих растений. С. животных (живчик, семенная нить, спермий) у всех позвоночных и большинства беспозвоночных имеет жгутик (хвост, рис., а), обеспечивающий его поступательное движение; у некоторых червей, многоножек, ракообразных и клещей С. не имеют хвоста и характеризуются большим разнообразием строения и амёбоидным движением. Длина С. у различных видов измеряется десятками и сотнями микрометров, у насекомых достигает нескольких миллиметров. Хвостатые С. имеют сравнительно короткую (округлую, конусовидную, крючковидную и т.д.) головку, в которой расположено ядро, содержащее конденсированный хроматин. На переднем конце головки находится акросома, содержащая лизины. За головкой следует более тонкий промежуточный отдел, переходящий в нитевидный хвост. Промежуточный отдел у С. большинства видов животных короткий и заключает в себе базальное тело (центриоль), состоящее из девяти расположенных кольцом триплетов коротких микротрубочек и окруженное венчиком из 4—10 крупных митохондрии. От базального тела берёт начало осевая нить (осевой комплекс, аксонема), сокращение которой обеспечивает биение жгутика и перемещение С. Ультраструктура осевой нити у разных С. практически одинакова и сходна с таковой у ресничек (рис., б, в); она состоит из фибрилл (микротрубочек, диаметром 200—250 Ǻ), 2 из которых лежат в центре, а 9 периферических двойных (дублеты) расположены кольцом вокруг центральных (9 + 2). Иногда снаружи от дуолетов лежат ещё 9 дополнительных фибрилл (9 + 9 + 2). Периферические фибриллы жгутика непосредственно переходят в микротрубочки базального тела, центральные фибриллы остаются свободными. В осевых нитях жгутиков (в фибриллах и структурах, связанных непосредственно с ними) имеются белки, подобные актину и миозину скелетных мышц и обладающие способностью расщеплять АТФ. У насекомых головка С. сильно вытянута в длину, иногда спирально закручена и без заметных границ переходит в длинный хвост. Большая часть хвоста содержит особую структуру митохондриального происхождения, называется побочным ядром. У многих животных в семенниках образуется несколько типов С., большинство которых атипичны и возникают в результате нарушения хода сперматогенеза, главным образом в период делений созревания; такие С., по-видимому, не обладают оплодотворяющей способностью. С. растений (антерозоиды) образуются в органах, называемых антеридиями. У большинства семенных растений С., обычно называют спермиями, лишены жгутиков и активно не двигаются (оплодотворение происходит не в жидкой среде). С. с двумя или многими жгутиками имеются у зелёных и бурых водорослей, у некоторых низших грибов, мхов, папоротников, хвощей, плаунов, изоэтесов, селагинелл, саговников и гинкго. С. лишены целлюлозной оболочки, в большинстве случаев очень мелкие (за исключением С. некоторых саговников, у которых они достигают 300 мкм в диаметре и видны невооружённым глазом), но с крупным ядром и небольшим количеством цитоплазмы. Жгутик находится на переднем (по направлению движения) конце С. Движение С. к яйцеклетке определяется различными химическими веществами, входящими в состав женских половых клеток и органов.
27. 1. Оплодотворение— слияние мужской и женской половых клеток, в результате чего восстанавливается диплоидный набор хромосом, характерный для данного вида животных, и возникает качественно новая клетка — зигота (оплодотворенная яйцеклетка, или одноклеточный зародыш)
2.Биологическое значение оплодотворения состоит в том, что при слиянии женской и мужской половых клеток образуется новый организм, несущий в себе признаки и матери и отца. При образовании половых клеток в мейозе возникают гаметы с разным сочетанием хромосом, поэтому образующиеся после оплодотворения новые организмы могут сочетать в себе признаки обоих родителей в самых различных комбинациях. В результате этого происходит увеличение наследственного разнообразия организмов.
3.Этапы оплодотворения:
В процессе оплодотворения различают три фазы: 1) дистантное взаимодействие и сближение гамет; 2) контактное взаимодействие и активизация яйцеклетки; 3) вхождение сперматозоида в яйцо и последующее слияние — сингамия. I стадия — дистантное взаимодействие, включает в себя 3 механизма:
хемотаксис — направленное движение сперматозидов навстречу к яйцеклетке (гинигамоны 1,2);
реотаксис — движение сперматозоидов в половых путях против тока жидкости;
капацитация — усиление двигательной активности сперматозоидов, под воздействием факторов женского организма (рН, слизь и другие).
II стадия — контактное взаимодействие, за 1,5-2 ч сперматозоиды приближаются к яйцеклетке, окружают ее и приводят к вращательным движениям, со скоростью 4 оборота в минуту. Одновременно из акросомы сперматозоидов выделяются сперматозилины, которые разрыхляют оболочки яйцеклетки. В том месте, где оболочка яйцеклетки истончается максимально, происходит оплодотворение, оволемма выпячивается, и головка сперматозоида проникает в цитоплазму яйцеклетки, занося с собой центриоли, но оставляя снаружи хвостик.
III стадия — проникновение, самый активный сперматозоид проникает головкой в яйцеклетку, сразу после этого в цитоплазме яйцеклетки образуется оболочка оплодотворения, которая препятствует полиспермии. Затем происходит слияние мужского и женского пронуклеусов, этот процесс носит название синкарион. Этот процесс (сингамия) и есть собственно оплодотворение, появляется диплоидная зигота (новый организм, пока одноклеточный).
Кортикальная реакция - слияние плазмолеммы яйцеклетки с мембранами кортикальных гранул, в результате чего содержимое из гранул выходит в перивителлиновое пространство и воздействует на молекулы гликопротеидов блестящей зоны. Образуется оболочка оплодотворения толщиной 50 нм, препятствующая полиспермии — проникновению других спермиев.
4. Условия, необходимые для оплодотворения:
концентрация сперматозоидов в эякуляте, не менее 60 млн в 1 мл;
проходимость женских половых путей;
нормальная температура тела женщины;
слабощелочная среда в женских половых путях.
28.
Партеногенез—одна из форм полового размножения организмов, прикоторой женские половые клетки (яйцеклетки) развиваются во взрослом организме без оплодотворения.
Андрогенез— развитие яйцеклетки с мужским ядром, привнесённым в неё спермием в процессе оплодотворения. Андрогенез наблюдается у отдельных видов животных (шелкопряд) и растений (табак, кукуруза) в тех случаях, когда материнское ядро погибает до оплодотворения, которое при этом является ложным, то есть женское и мужское ядра не сливаютсяи в дроблении участвует только мужское ядро.
Гиногенез — частный случай партеногенеза, особая форма полового размножения, при которой после проникновения спермия в яйцеклетку их ядра не сливаются, и в последующем развитии участвует только ядро яйцеклетки, либо не происходит оплодотворения. При этом нет объединения наследственного материала родителей посредством слияния ядер их половых клеток. Роль сперматозоида ограничивается активацией осеменённого яйца к развитию. В природе гиногенез встречается крайне редко. Известен у нескольких видов рыб (голомянка, серебряный карась и др.), земноводных, круглых червей и растения семейства амариллисовых.
29.
1. Прогамный. Пол будущего организма определяется в ходе гаметогенеза у родительских
особей.
2. Сингамный. Пол будущего организма определяется в момент слияния половых клеток.
3. Эпигамный. Пол будущего организма определяется в процессе онтогенеза. У человека
имеет место переопределение пола (при патологии) – хотя истинного нет. |
|
|
 Скачать 154.98 Kb.
Скачать 154.98 Kb.