биологическая химия 53. 1. Строение и классификация протеиногенных аминокислот
 Скачать 176 Kb. Скачать 176 Kb.
|

1. Строение и классификация протеиногенных аминокислотПротеиногенными называются 20 аминокислот, которые кодируются генетическим кодом и включаются в белки в процессе трансляции. Классификация этих аминокислот основана как на строении, так и на полярности боковых цепей. В таблице для каждой из аминокислот приводятся следующие характеристики: — классификация (семь классов); — название и принятое сокращение (по трем первым буквам) (например, для гистидина — Нis); — однобуквенный символ, удобный при записи белковых последовательностей (для гистидина — Н); — полярность боковой цепи (для гистидина +10,3): чем выше эта величина, тем более полярна молекула аминокислоты. На схеме по мере увеличения полярности окраска поля с названием аминокислоты меняется от желтых тонов через зеленые к синим. Для ионогенных групп боковой цепи приведены рКa (цифры красного цвета). К алифатическим аминокислотам относятся глицин, аланин, валин, лейцин и изолейцин. Эти аминокислоты не несут в боковой цепи гетероатомов (N, О или S), циклических группировок и характеризуются отчетливо выраженной низкой полярностью. Также малополярны серосодержащие аминокислоты— метионин и цистеин, причем цистеин существует лишь в недиссоциированном состоянии. Благодаря образованию дисульфидных мостиков, цистеин выполняет важную функцию стабилизации пространственной структуры белков. Аминокислота цистин состоит из двух остатков цистеина, соединенных дисульфидным мостиком. Ароматические аминокислоты содержат мезомерные (резонансно стабилизированные) циклы. В этой группе лишь фенилаланин проявляет низкую полярность. Тирозин и триптофан характеризуются заметной, а гистидин — даже высокой полярностью. Имидазольное кольцо гистидина заметно протонируется уже при слабокислых значениях рН. Поэтому гистидин, обладающий ароматическими свойствами лишь в протонированной форме, может быть отнесен к основным аминокислотам. Тирозин и триптофан сильно поглощают в УФ-области спектра между 250 и 300 нм. Нейтральные аминокислоты содержат гидроксильные (серин, треонин) или карбоксамидные группы (аспарагин, глутамин). Хотя амидные группы неионогенны, молекулы аспарагина и глутамина высоко полярны. Карбоксильные группы боковых цепей кислых аминокислот — аспарагиновой и глутаминовой — полностью ионизированы во всем диапазоне физиологических значений рН. Аналогичным образом, боковые цепи основных аминокислот — лизина и аргинина — полностью протонированы в нейтральной области рН. Сильно основной, а, следовательно, очень полярной аминокислотой, является аргинин, содержащий гуанидиновую группировку. Особое положение занимает пролин. Боковая цепь пролина состоит из пятичленного цикла, включающего α-углеродный атом и α-аминогруппу. Поэтому пролин, строго говоря, является не амино-, а иминокислотой. Атом азота и кольце является слабым основанием и не протонируется при физиологических значениях рН. Благодаря циклической структуре пролин вызывает изгибы полипептидной цепи, что очень существенно для структуры коллагена. Некоторые из перечисленных аминокислот не могут синтезироваться в организме человека и должны поступать вместе с пищей. Эти незаменимые аминокислоты отмечены звездочками красного цвета.  2. Механизм действия ферментов. Теория промежуточных соединенийФермент взаимодействует с субстратом и образует короткоживущий фермент-субстратный комплекс. По завершении реакции, фермент-субстратный комплекс распадается на продукты и фермент. Фермент в итоге не изменяется: по окончании реакции он остается таким же, каким был до неё, и может теперь взаимодействовать с новой молекулой субстрата (рис. 1). 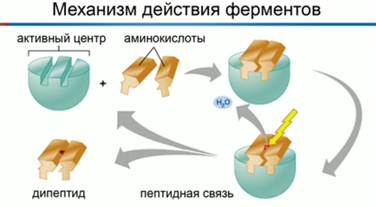 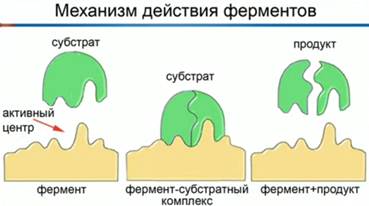 Рис. 1. Механизм взаимодействия фермента и субстрата На рисунке 3 представлен механизм работы фермента, в частности, образования пептидной связи между молекулами аминокислот. Две аминокислоты взаимодействуют между собой в активном центре фермента, между ними образуется пептидная связь. Новое вещество (дипептид) покидает активный центр фермента, поскольку оно по своей структуре не соответствует этому центру. Особенностью ферментов является то, что они обладают высокой специфичностью, т. е. могут ускорять только одну реакцию или реакции одного типа. В 1890 году Э. Г. Фишер предположил, что эта специфичность обусловлена особой формой молекулы фермента, которая точно соответствует форме молекулы субстрата. Эта гипотеза получила название «ключа и замка», где ключ сравнивается с субстратом, а замок – с ферментом. Гипотеза гласит: субстрат подходит к ферменту, как ключ подходит к замку. Избирательность действия фермента связана со строением его активного центра (рис. 2). 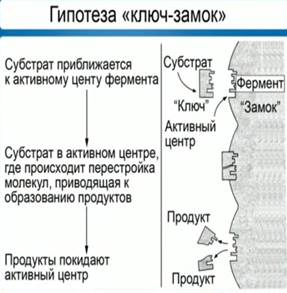 Рис. 2. Гипотеза взаимодействия фермента и субстрата по принципу ключ-замок Э. Г. Фишера В первую очередь, на активность фермента влияет температура. С повышением температуры скорость химической реакции возрастает. Увеличивается скорость молекул, у них появляется больше шансов столкнуться друг с другом. Следовательно, увеличивается вероятность того, что реакция между ними произойдет. Температура, обеспечивающая наибольшую активность фермента – оптимальная. За пределами оптимальной температуры скорость реакции снижается вследствие денатурации белков. Когда температура снижается, скорость химической реакции тоже падает. В тот момент, когда температура достигает точки замерзания, фермент инактивируется, но при этом не денатурирует (см. видео). В наше время для длительного хранения продуктов широко используют способ быстрого замораживания. Оно останавливает рост и развитие микроорганизмов, а также инактивирует ферменты, находящиеся внутри микроорганизмов, и предотвращает разложение продуктов питания. Кроме этого, активность ферментов зависит ещё от pH среды (кислотности – то есть показателя концентрации ионов водорода). В большинстве случаев, ферменты работают при нейтральном pH, т. е. при pH около 7. Но существуют ферменты, которые работают либо в кислой и сильнокислой, либо в щелочной и сильнощелочной среде. Например, один из таких ферментов – пепсин, он находится у нас с вами в желудке, работает в сильнокислой среде и расщепляет белки. Поскольку в желудке среда достаточно кислая, 1,5 – 2 pH, то этот фермент работает при сильнокислой среде. Ферменты подвержены действию активаторов и ингибиторов. Некоторые ионы, например, ионы металлов Mg, Mn, Zn активируют ферменты. Другие же ионы (к ним относятся ионы тяжелых металлов, а именно Hg, Pb, Cd), наоборот, подавляют активность ферментов, денатурируют их белки. Выдвинутая в 1913 году Л. Михаэлисом и М. Ментен общая теория ферментативного катализа постулировала, что фермент Е сначала обратимо и относительно быстро связывается с со своим субстратом S в реакции: E + S = ES Образовавшийся при этом фермент-субстратный комплекс ES, не имеющий аналогий в органической химии и химическом катализе, затем распадается в второй более медленной (лимитирующей) стадии реакции: ES = Е + Р Этапы взаимодействия фермента и субстрата1 этап: происходит сближение и ориентация субстрата относительно субстратного центра фермента и его постепенное «причаливание» к «якорной» площадке. 2 этап: напряжение и деформация: индуцированное соответствие - происходит присоединение субстрата, которое вызывает конформационные изменения в молекуле фермента приводящие к напряжению структуры активного центра и деформации связанного субстрата. 3 этап: непосредственный катализ. Имеется реакция: S → P + Q Представим эту реакцию в виде отдельных новых стадий: S + E = ES = E + P подстадии:
4.ES** = ES*** 5.ES*** = EP 6.EP = E + P 3. Водорастворимые витамины, условия развития гиповитаминозов, коферментная роль. Примеры |