|
|
цикл кребса. 1я реакция образование лимонной кислоты
Этот метаболический путь назван именем открывшего его автора – Г. Кребса, получившего (совместно с Ф. Липманом) за данное открытие в 1953 г. Нобелевскую премию. В цикле лимонной кислоты улавливается большая часть свободной энергии, образующейся при распаде белков, жиров и углеводов пищи. Цикл Кребса – центральный путь обмена веществ.
Образовавшийся в результате окислительного декарбоксилирования пирувата ацетил-КоА в матриксе митохондрий включается в цепь последовательных реакций окисления. Таких реакций восемь.
1-я реакция – образование лимонной кислоты. Образование цитрата происходит путем конденсации ацетильного остатка ацетил-КоА с оксалацетатом (ОА) при помощи фермента цитратсинтазы (с участием воды):
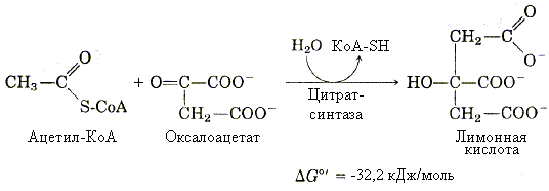
Данная реакция практически необратима, поскольку при этом распадается богатая энергией тиоэфирная связь ацетилS-КоА.
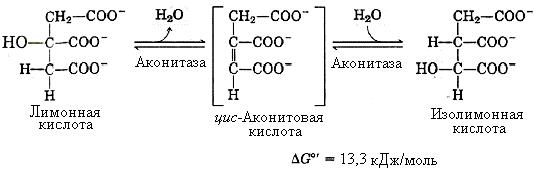
2-я реакция – образование изолимонной кислоты. Эта реакция катализируется железосодержащим (Fe – негеминовое) ферментом – аконитазой. Реакция протекает через стадию образования цис-аконитовой кислоты (лимонная кислота подвергается дегидратации с образованием цис-аконитовой кислоты, которая, присоединяя молекулу воды, превращается в изолимонную).
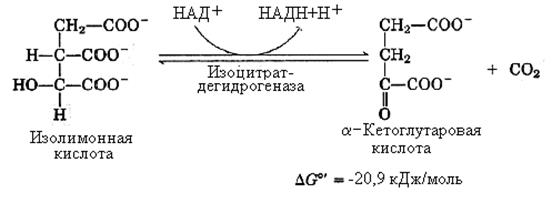
3-я реакция – дегидрирование и прямое декарбоксилирование изолимонной кислоты. Реакция катализируется НАД+–зависимым ферментом изоцитратдегидрогеназой. Фермент нуждается в присутствии ионов марганца (или магния). Являясь по своей природе аллостерическим белком, изоцитратдегидрогеназа нуждается в специфическом активаторе – АДФ.
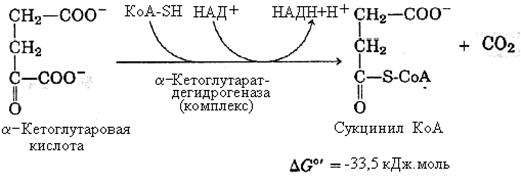
4-я реакция – окислительное декарбоксилирование α-кетоглутаровой кислоты. Процесс катализируется α-кетоглутаратдегидрогеназой – ферментным комплексом, по структуре и механизму действия похожим на пируватдегидрогеназный комплекс. В его состав входят те же коферменты: ТПФ, ЛК и ФАД – собственные коферменты комплекса; КоА-SH и НАД+ – внешние коферменты.
5-я реакция – субстратное фосфорилирование. Суть реакции заключается в переносе богатой энергией связи сукцинил-КоА (макроэргическое соединение) на ГДФ с участием фосфорной кислоты – при этом образуется ГТФ, молекула которого вступает в реакцию перефосфорилирования с АДФ – образуется АТФ.
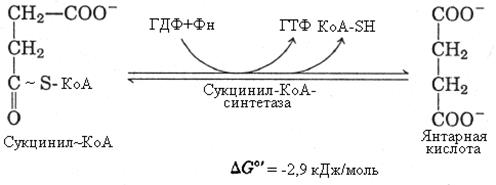
6-я реакция – дегидрирование янтарной кислоты сукцинатдегидрогеназой. Фермент осуществляет прямой перенос водорода с субстрата (сукцината) на убихинон внутренней мембраны митохондрий. Сукцинатдегидрогеназа - II комплекс дыхательной цепи митохондрий. Коферментом в этой реакции является ФАД.
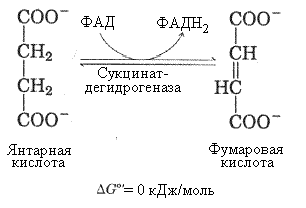
7-я реакция – образование яблочной кислоты ферментом фумаразой. Фумараза (фумаратгидратаза) гидратирует фумаровую кислоту – при этом образуется яблочная кислота, причем ее L-форма, так как фермент обладает стереоспецифичностью.
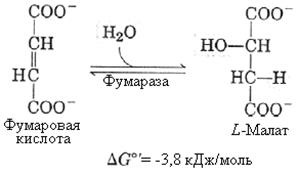
8-я реакция – образование оксалацетата. Реакция катализируется малатдегидрогеназой, коферментом которой служит НАД+. Образовавшийся под действием фермента оксалацетат вновь включается в цикл Кребса и весь циклический процесс повторяется.
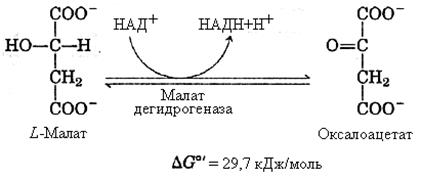
Последние три реакции обратимы, но поскольку НАДН?Н+ захватывается дыхательной цепью, равновесие реакции сдвигается вправо, т.е. в сторону образования оксалацетата. Как видно, за один оборот цикла происходит полное окисление, “сгорание”, молекулы ацетил-КоА. В ходе цикла образуются восстановленные формы никотинамидных и флавиновых коферментов, которые окисляются в дыхательной цепи митохондрий. Таким образом, цикл Кребса находится в тесной взаимосвязи с процессом клеточного дыхания.
Функции цикла трикарбоновых кислот многообразны:
· Интегративная – цикл Кребса является центральным метаболическим путем, объединяющим процессы распада и синтеза важнейших компонентов клетки.
· Анаболическая – субстраты цикла используются для синтеза многих других соединений: оксалацетат используется для синтеза глюкозы (глюконеогенез) и синтеза аспарагиновой кислоты, ацетил-КоА – для синтеза гема, α-кетоглутарат – для синтеза глютаминовой кислоты, ацетил-КоА – для синтеза жирных кислот, холестерола, стероидных гормонов, ацетоновых тел и др.
· Катаболическая – в этом цикле завершают свой путь продукты распада глюкозы, жирных кислот, кетогенных аминокислот – все они превращаются в ацетил-КоА; глутаминовая кислота – в α-кетоглутаровую; аспарагиновая – в оксалоацетат и пр.
· Собственно энергетическая – одна из реакций цикла (распад сукцинил-КоА) является реакцией субстратного фосфорилирования. В ходе этой реакции образуется одна молекула ГТФ (реакция перефосфорилирования приводит к образованию АТФ).
· Водороддонорная – при участии трех НАД+-зависимых дегидрогеназ ( дегидрогеназ изоцитрата, α-кетоглутарата и малата) и ФАД-зависимой сукцинатдегидрогеназы образуются 3 НАДН?Н+ и 1 ФАДН2. Эти восстановленные коферменты являются донорами водорода для дыхательной цепи митохондрий, энергия переноса водородов используется для синтеза АТФ.
· Анаплеротическая – восполняющая. Значительные количества субстратов цикла Кребса используются для синтеза разных соединений и покидают цикл. Одной из реакций, восполняющих эти потери, является реакция, катализируемая пируваткарбоксилазой.
Скорость реакция цикла Кребса определяется энергетическими потребностями клетки
Скорость реакций цикла Кребса коррелирует с интенсивностью процесса тканевого дыхания и связанного с ним окислительного фосфорилирования – дыхательный контроль. Все метаболиты, отражающие достаточное обеспечение клетки энергией являются ингибиторами цикла Кребса. Увеличение соотношения АТФ/АДФ – показатель достаточного энергообеспечении клетки и снижает активность цикла. Увеличение соотношения НАД+/ НАДН, ФАД/ ФАДН2 указывает на энергодефицит и является сигналом ускорения процессов окисления в цикле Кребса.
Основное действие регуляторов направлено на активность трех ключевых ферментов: цитратсинтазы, изоцитратдегидрогеназы и a-кетоглутаратдегидрогеназы. Аллостерическими ингибиторами цитратсинтазы являются АТФ, жирные кислоты. В некоторых клетках роль ее ингибиторов играют цитрат и НАДН. Изоцитратдегидрогеназа аллостерически активируется АДФ и ингибируется при повышении уровня НАДН+Н+.
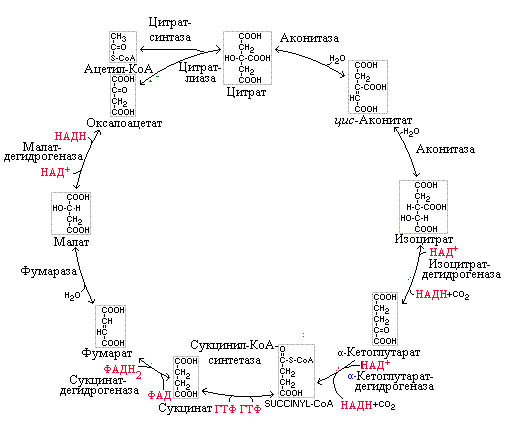
Рис. 5.15. Цикл трикарбоновых кислот (цикл Кребса)
Последний является ингибитором и a-кетоглутаратдегидрогена зы, активность которой снижается также при повышении уровня сукцинил-КоА.
Активность цикла Кребса во многом зависит от обеспеченности субстратами. Постоянная “утечка” субстратов из цикла (например, при аммиачном отравлении) может вызывать значительные нарушения энергообеспеченности клеток. |
|
|
 Скачать 116.82 Kb.
Скачать 116.82 Kb.