горомны биохимичя. Биохимия гормонов
 Скачать 0.56 Mb. Скачать 0.56 Mb.
|

|
Биохимия гормонов. Биологические мембраны, их основные функции. Жидкостно-мозаичная модель биологической мембраны. Общие принципы построения мембран одинаковы. Мембраны отделяют клетку от окружающей среды и делят клетку на отсеки. Мембраны регулирует транспорт веществ клетки, обеспечивают специфику межклеточных контактов и принимают сигналы из внешней среды. Основу мембрана составляет билипидный слой, в структуру которого включены белки, также в составе мембраны содержатся углеводы (до 1%). Липидный бислой, его функции. Классификация и роль белков в структуре мембраны. Липидный бислой образует 2 рядами амфифильных молекул, гидрофобный компоненты которых направлены внутрь, а гидрофильные группы направлены наружу и контактируют с внешний средой. Основные липиды мембран – это фосфолипиды (фосфатидилиэтанолин, фосфатидилсерин, фосфатидихолин), с 2ух углеродным атомом данных фосфолипидов связан остаток полиненасыщенных жирных кислот. Холестерол присутствует в мембранах всех клеток, придает мембранам жесткость и снижает их тягучесть. Молекула холестерола располагается в гидрофобном участки мембраны и соотношение холестерола и других липидов в мембране приблизительно 0,3:0,9 Наибольшее содержание холестерола в цитоплазматической мембране. Гликолипиды - это комплекс липида с углеводом. Углеводные компоненты чаще всего направлены во внешнюю среду и выполняют рецепторную функцию. В целом липиды выполняют функцию: 1) формулируют бислой; 2) создают среду для функционирования мембранных белков; 3) регулируют активность ферментов; 4) участвуют в передаче сигнала. Белки мембраны по расположению подразделяются на: Интегральные Полность. Пронизывают билипидный слов и являются амфифильными Полуинтегральные Наполовину пронизывают билипидный слой Периферические Локализованы либо снаружи, либо с внутренней стороны. Белки, которые находятся на наружных поверхностях мембраны, часто гликолизированы, то есть содержат углеводный компонент, что защищает их от действия протеас. Функции белков: 1) обеспечивает избирательный транспорт веществ; 2) участвуют в передаче гормонального сигнала; 3) участвуют в образовании окаймленных ямок, необходимых для эндо- и экзоцитоза; 4) являются ферментами; 5) участвуют в реакциях иммунитета; 6) обеспечивают межклеточные контакты. 3 Транспорт веществ через биологическую мембрану: пассивный и активный, виды, различия между ними. Транспорт ионов органических молекул через мембрану может проходить по градиенту концентрации – это пассивный транспорт. Против градиента концентрации - это активный транспорт. Пассивный транспорт может осуществляться следующими способами: Простая диффузия - это движение веществ по градиенту концентрации, без участия белков переносчиков, таким образом, транспортируется: кислород, вода, углекислый газ, мочевина, спирт и другие гидрофобный низкомолекулярные соединения. Облегченная диффузия - происходит по градиенту концентрации при участии белков переносчиков. Например: таким образом, транспортируется глюкоза при участии глюкозных транспортеров. Пассивный симпорт - это перенос двух ионов по градиенту концентрации в одном направлении. Например: транспорт ионов водорода и фосфат ионов Пассивный антипорт - это перенос ионов по градиенту концентрации в противоположном направлении. Например: транспорт аминов хлора и гидрокарбонат ионов. Активный транспорт: Первично-активный транспорт Происходит против градиента концентрации с затратой АТФ при участии транспортных АТФ-фазы. Например: протонная АТФ-аза, которая обеспечивает кислую среду клетки в лизосомах; кольцевая АТФ-аза обеспечивает низкую концентрацию кальция в цитозоли и создает депо кальция в эндоплазматическом ретикуле и митохондриях. Натрийкаливая АТФ-аза обеспечивает формирование мембранного потенциала клетки. Вторично-активный транспорт. Происходит за счет градиента концентрации одного из переносимых веществ, который чаще всего создается натрийкалиевой АТФ-азой, функционирующая с затратой АТФ. Чаще всего "активный симпорт и антипорт" А. Активный симпорт - Б. Активный антипорт - перенос двух веществ одновременно в одном направлении, одно из которых перемещается против градиента концентрации, за счет перемещения другого вещества по градиенту концентрации. Например: Na-зависимый транспорт кальция клетки слюнных желез. Эндо- и экзоцитоз. Перенос веществ из внеклеточной среды в клетку макромолекул (белки, нуклеиновые кислоты, более крупные частицы) происходит путем эндоцитоза. Макромолекула связывается в определенных участках мембраны, которые называются окаймленными ямками. Эндоцитоз происходит при помощи рецепторов встроенных в окаймленные ямки, что позволяет клеткам поглощать специфические вещества. Макромолекулы (гормоны, ферменты, липопротеины) синтезируются в кровь или межклеточное пространство путем экзоцитоза. Чаще всего экзоцитоз регулируется путем изменения концентрации ионов Ca в цитоплазме клетки. Понятие о "гормонах", "эндокринных железах. Отличие эндокринных желез от экзокринных. Свойства гормонов. Гормоны - биологически активные вещества, действующие как химические посредники, переносящие информацию об изменениях условий внешней и внутренней среды в различные органы и ткани. Гормоны секретируется эндокринными железами, выделяемыми гормоны в кровь (венозную) или лимфу. Однако, есть органы, которые продуцируют гормоны. Сердце продуцирует натрийуретический пептид Почки продуцируют ренин, эритропоэтин Печень продуцирует ИФР (инсулиноподобный фактор роста) Жировая ткань продуцирует лептин Желудочно - кишечный тракт продуцирует холецистокинин (хцк), гастрин, грелин Тромбоциты продуцирует фактор роста тромбоцитов (тфр) Свойства гормонов: Обладают дистантным действием, то есть действует вдали от места синтеза. Действует в низких концентрациях их действия обычно кратковременно. Например, период распада для катехоламинов 2 - 3 минуты, тироксина - 12 ч, стероидов - от 4 минут до 2 часов, полипептидов 4 - 40 минут, белков от 15 минут до 3 ч. Специфичность Клетки, на которые действует гормон - мишени, на них локализованы рецепторы к данному гормону. Классификация гормонов по химическому строению, биологическим функциям, по механизму действия. Классификация гормонов по химической природе: Производные аминокислот Гормоны мозгового слоя надпочечников Дофамин Катехоламины (норадреналин и адреналин) Йодсодержащие гормоны щитовидной железы (трийодтиронин-т3 и тироксин-т4) Катехоламины и йодсодержащего гормоны являются производными аминокислоты тирозина. Пептиды и белки Могут состоять из небольшого количества аминокислот или содержать большое количество аминокислотных остатков. Основная масса гормонов являются белками или полипептидами. Стероидные гормоны Являются производными холистерола Половые гормоны Гормоны коры надпочечников (глюкокортикоиды и минералкортикоиды) Функции гормонов: Влияют на репродуктивные процессы Гормоны продуцируемые половыми железами и передней доли гипофиза. Они регулируют рост и структурную целостность репродуктивных органов, половое поведение, фенотипическое различие между полами и продолжение рода. Влияют на рост и развитие Классическими гормонами, участвующими в процессе роста, являются соматотропный гормон (СТГ), гормоны роста (ГР), тиреоидные гормоны, инсулин, глюкокортикоиды, половые гормоны - стимулирующие стг на рост тела опосредуется семейством пептидов, называемых инсулиноподобные факторы роста (ИФР0. Тиреоидные гормоны, особенно Т4, играют роль в индукции роста и клеточной дифференцировке. Эффект Т4 в мозге опосредуется продуцированием фактора роста нервов (фрн).Поддержание внутренней среды-гомеостаз Принимает участие множество гормонов. Например, вазопрессин и альдостерон влияют на водно - солевой обмен и регуляцию артериального давления, объем внеклеточной жидкости и электролитические состав. Инсулин и глюкагон обеспечивают поддержание постоянство глюкозы в крови. Регуляции энергетического обмена Центральную роль играет гипоталамус, который интегрирует афферентные сигналы и способствуют возникновению эфферентных сигналов, регулировать пищевое поведение и энергозатраты. В регуляции энергетического обмена участвуют инсулин, глюкоидные и тиреоидные гормоны. Эффекты, наблюдаемые в клетках мишенях гормонов: Изменяют проницаемость в клеточных мембранах для определенных веществ Меняют активность уже существующих ферментов или белков Меняют количество ферментов или белков По механизму действия гормоны делят на 2 класса: Гормоны с мембранным механизмом действия К ним относят: белково-пептидные гормоны, катехоламины Какие гормоны не проникают через клеточную мембрану. Рецепторы расположенные на поверхности мембраны, а информация о гормональном сигнале передается через систему вторичных посредников. Гормоны с мембранным механизмом действия чаще всего меняют активность уже существующих ферментов или белков. Гормоны внутриклеточным (цитозольным) механизмом действия Стероидные и тиреоидные гормоны. Такие гормоны легко проникают через клеточную мембрану и связываются с рецептором в цитозоль или ядре клетки. Они способствуют синтезу новых ферментов или белков. Аденилатциклазый, инозитолфосфатный, цитозольный механизм действия гормонов. Изменения, возникающие в клетках мишенях при действии гормонов. Аденилатциклазная система (АЦ - система) Включает в себя рецептор первичного мессенджера активатора АЦ-системы, рецептор ингибитора АЦ-системы, фермент аденилатциклазу (АЦ). G-белок, который состоит из трёх субъединиц: альфа, бета и гамма. Альфа-субъединица связана с молекулой ГДФ. Цитозольный фермент - протеинкиназа А (ПКА). Последовательность событий передачи сигнала с помощью АЦ-системы. Гормон связывается с рецептором на поверхности клеточной мембраны с образованием гормон рецепторного комплекса, который активирует встроенную мембраны G-белок. Активация G-белка способствует диссоциации белка на альфа-субъединицу. 2 единица включает: бета и гамма субъединицу. При активации G-белка молекула ГДФ структуре Альфа субъединицы заменяется на ГТФ и комплекс альфа-субъединицы заменяется на ГТФ, активирует встроенный в мембрану фермент аденилатциклазу, которые катализируют реакцию АТФ в цАМФ. Образовавшийся вторичный посредник циклический АМФ поступает в цитозоль клетки и активируют протеинкиназуА, которая состоит из 4 субъединиц:2 регуляторных и 2 каталитических. Активация протеинкиназыА приводит к диссоциации регуляторных и каталитических субъединиц. Регуляторные субъединицы ферментативной активностью не обладают, а каталитические при участии молекулы АТФ фосфорилируют белки, ферменты по остаткам серина, трианина, что повышает или понижает их активность и соответственно изменяется скорость метаболических процессов в клетке. Инактивация аденилатциклазы происходит при участии фермента фосфодиэстеразы, который переводит циклический АМФ в АМФ. 2. Инозитолфосфатная система Включает в себя рецепторы (интегральный белок), фермент фосфолипазу С, G-белок, а также белки кальмодулин, протеинкиназу С (ПКС). Последствия механизмов Гормон связывается с рецептором на поверхности клеточной мембраны, в результате образуется гормонрецепторный комплекс, который активируют встроенный в мембрану G-белок. Комплекс альфа-субъединицы +гтф активирует мембранные фермент фосфолипазу С, который обеспечивает гидролиз фосфолипида клеточной мембраны - фосфатидилинозитол-4,5-дифосфата (ФИФ2). В ходе реакции образуются 2 вторичных посредника: диацилглицерол (ДАГ), который остаётся в мембране и участвует в активации протеинкиназыС и образуются инозитол-3-фосфат, который является гидрофильным и уходит в цитозоль. Инозитол-3-фосфат связывается с Ca-каналами эндоплазматического ретикулума, в результате кальций поступает в цитозоль клетки и связывается с кальций связывающим белком - кальмодулином. Кальцикальмодулиновый комплекс активирует протеинкиназуС, которая состоит из 2 субъединиц: регуляторной и каталитической. Активация протеинкиназыС приводит к диссоциации субъединиц: каталитическая субъединица катализируют фосфорилирование белков, ферментов с изменением скорости метаболизма. Поступивший в цитозоль клетки кальций может самостоятельно менять активность белков и ферментов. На диацилглицерол может действовать фермент - диациглицероллипаза, который катализирует гидролиз сложной эфирной связи во втором положении где локализованы полиненасыщенные жирные кислоты, в результате образуются свободные жирные кислоты, которые являются источниками эйкозаноидов. Действие инозитолфосфатной системы не продолжительно, так как повышение концентрации кальция в инозитоле активирует кальциевую АТФ фазу, которая выкачивает ионы кальция из цитозоля в эндоплазматический ретикулум. Эффекты наблюдаемые в клетках мишенях под влиянием гормонов: 1. Изменяется проницаемость клеточной мембраны для определённых веществ 2. Меняется активность уже существующих ферментов или белков 3. Меняется количество ферментов (белков) Роль гипоталамуса в регуляции эндокринных функций. Структура и функция либеринов и статинов. Гипоталамус- это высший нейроэндокринный орган, в котором происходит интеграция нервной и эндокринной систем. В гипоталамусе вегетативная нервная система оказывает регулирующие влияние на эндокринную систему, а та регулирующие деятельность нервной системы. В результате формируется нейро-эндокринная система, регулирующая все физиологические функции. В гипоталамусе синтезируются релизин факторы (РФ), которые оказывают воздействие на переднюю долю гипофиза, на выработку тропных гормонов РФ делят на 2 типа: 1. Либерины (усиливают выработку тропных гормонов гипофиза). 2. Статины ( обладают противоположным действием). В настоящее время выделяют: Соматолиберин стимулирует выработку соматотропного гормона (СТГ). Тиреолиберин стимулирует выработку тиркотропного гормона (ТТГ). Кортиколиберин стимулирует выработку кортикотропного гормона (АКТГ). Пролактолиберин стимулирует выработку пролактина. Миланолиберин стимулирует выработку миланотропного гормона. Фоллиберин стимулирует выработку фоликулостимулирующего гормона. Люлиберин стимулирует выработку лютеинозирующего гормона (ЛГ). Фоллиберин и Люлиберин – это ганадолиберин. Соматостатин тормозит секрецию СТГ и ТТГ. Соматостатин, кроме гипоталамуса, синтезируется в D-клетках островов Лангерганса поджелудочной железы и в кишечнике. Пролактостатин тормозит секрецию пролактин. Миланостатин подавляет секрецию миланотропного гормона. В гипоталамусе синтезируются 2 гормона – окситацин и вазопрессин, которые по аксонам нейронов поступают в заднюю долю гипофиза, где хранятся и откуда высвобождаются. Гормоны передней доли гипофиза (СТГ, ЛТГ, ФСГ, ЛТ, ТТГ, АКТГ, пролактин): химическая природа, биологическая роль. Принцип обратной связи на примере регуляции секреции ТТГ и АКТГ. В передней доле гипофиза (аденогипофиз) выделяют клетки, которые синтезируют тропные гормоны. Тропные гормоны оказывают влияние на периферические эндокринные железы (- ганадотропы синтезируют ФСГ И ЛГ). На долю этих клеток приходится 10-15% клеточного состава аденогипофиза. Тиреотропы, в них синтезируется ТТГ, на их долю приходится ≈ 10% клеток аденогипофиза. Соматотррпы, в них синтезируются СТГ, на их долю приходится до 50% клеток аденогипофиза. Кортикотропы , в них синтезируются АКТГ, на их долю приходится 15-20% клеток аденогипофиза. Лактотропы обеспечивают синтез пролактин а, составляет 10-25% клеток аденогипофиза. Гормоны: ТТГ. Является гликопротеином, состоит из альфа- и бетта-субъединиц. Альфа-субъединица у всех одинаковая. ТТГ стимулирует функции щитовидной железы, поступление неорганического йода в щитовидную железу, синтез стереоглобулина, йодирование тирощина, рост щитовидной железы и её кровоснабжение. АКТГ (кортикотпопин) .Является пептидом, стимулирующий функцию коры надпочечников,её рост, кровоснабжение и синтез кортизола. А также альдестерола (уровень регулирующий ренинантеотензиновой системой и содержащей кальций и натрий в сыворотке крови). Ганадотропные гормоны.К ним относятся: фоликулосинтезирующие (ФСГ) и лютеинозирующие. Являются гликопротеинами, состоят из альфа- и бетта-субъединиц. Альфа-субъединица у всех одинаковая, а бетта-субъединица различна и определяет их биологическую активность. ФСГ стимулирует рост и созревание фолликулов , секрецию эстрогенов, стимулирует сперматогенез. ЛГ стимулирует синтез тестостерона, вызывает овуляцию, образует жёлтое тело, синтез и секрецию пролактина. Пролактин Стимулирует образование и секрецию молока, угнетает выработку тестостерона и сперматогенез. СТГ (гормон роста) Является полипептидом , в отличии от других гипофизарных гормонов не имеет эффекторной железы, мишенью являются все клетки организма. Функция гормона – обеспечение линейного роста и анаболическое действие на ткани. Действие на рост гормон оказывает не прямо, а через инсулиноподобный фактор роста (ИФР). Он стимулирует рост хрящей, костей, увеличивает синтез белка. Раньше назывался соматомедийным С. ИФР синтезируется в печени. СТГ в жировой ткани уменьшает усвоение глюкозы и усиливает липолиз (распад липидов). В мышцах снижает усвоение глюкозы, увеличивает освоение аминокислот, синтез белков и увеличивает массу тела за счёт белков. В печени усиливается глюконеогенез (синтез глюкозы из соединений неуглеводной природы и синтез ИФР). В крови гормон циркулирует с соматотропин-связывающим белком. Гормоны задней доли гипофиза. Место их синтеза, строение, физиологические эффекты. В ней находится вазопрессин (антидиоритический гомон (ФДГ)) и окситоцин. Синтезируется в гипоталамусе, хранятся в задней доле. Являются пептидами, имеющие в составе дисульфидные связи. Окситоцин Стимулирует сокращение гладкой мускулатуры матки при родах, стимулирует отделение молока путём сокращения миоэпителиальных клеток, окружающих альвеолы протоков молочной железы. Действует на сосуды и почки, его действие напоминает вазопрессин, снижает солевой аппетит. Вазопрессин Регулирует водно- солевой обмен, мишенью для него являются сосуды и почки. Действуя на почки, гормон повышает проницаемость собирательных трубочек для воды. Действуя на сосуды, гормон вызывает их сокращение. Конечным результатом действия гормонов является повышение остеомаляция мочи, снижение её объёма и снижение сосудов. Гормон, действуя на ЦНС , снижает температуру тела, облегчает консаледацию памяти. Гормоны щитовидной железы. Химическая природа. Физиологические и биохимические эффекты тиреоидных гормонов. Механизм влияния тироксина на скорость биологического окисления. В фолликулярных клетках синтезируются тиреоидные гормоны: 3-йодтиранин (Т3) и тироксин (Т4). Эти гормоны являются производными аминокислоты тирозина. Данные гормоны обладают цитозольным механизмом действия, в парах фолликулярных клеток С -клетках, синтезируется кальцитонин, который контролирует кальций фосфорный обмен, является полипептидом и обладает мембранным механизмом действия. В тиреоидных клетках синтезируется белок тиреоглобулин, который путем эндоцитоза поступает в полость “фолликула”, где осуществляется синтез Т3 и Т4. Для синтеза гормонов необходим неорганический, который поступает с водой и пищей. Гормоны, синтезируются из аминокислот: тирозин путем ее йодирования фенольного кольца, йод поступает путем активного транспорта при участии натрий калиевой АТФ-азы. Синтезируемые Т3 и Т4 поступают в кровоток и связываются с транспортными белками. Т4 практически весь связан с белком, и только 0,04% Т4 находится в свободном биологически активном состоянии. Белок транспортируемый Т4, называется тироксинсвязывающим белком. Он является глобулином, связывает до 15% Т4, остальная доля гормоны связывается с альбуминами и преальбуминами. Т3 в свободном состоянии 4%, остальная часть связана с тироксинсвязывающим глобулином или альбумином. Т3 и Т4 влияет на многие обменные процессы. В физиологической концентрации проявляют выраженное анаболическое действие, укоряет синтез белков, стимулирует рост и клеточную диффузию, повышает поглощение кислорода тканями организма, активирует работу натрий калиевой АТФазы, регулирует обмен веществ (особенно холистерола), углеводов. В высоких концентрациях Т3 и Т4 тормозят синтез белков, стимулирует катаболизм, в результате возникает азотный баланс, усиливается разобщение между фосфорилирование и окислением. Мишенью для кальцитонина являются кости и почки. Гормон понижает концентрацию кальция в крови, способствует транспорту кальция и фосфора из крови в костную ткань, подавляет реабсорбцию кальция в почках и тем самым стимулирует выведение кальция с мочой. Является антагонистом паратгормона. Скорость секреции кальцитонина зависит от уровня эстрогена, при недостатке эстрогена, секреты кальцитонина уменьшаются. Гормоны паращитовидных желез. Химическая природа. Биологическая роль. В них частично образуется кальцитонин. Паратгормон - полипептид, который повышает концентрацию Ca, снижает концентрацию фосфора в крови. Мишень - кости и почки. В костях активирует фермент щелочную фосфатазу и коллагеназу, которые действуют н костный матрикс вызывая его распад и мобилизацию Сa и фосфатов из кости в кровь. В почках стимулирует реабсорбцию Ca и тем самым задерживает его в организме и уменьшает реабсорбцию фосфатов. Строение, биосинтез и механизм действия кальцитриола (витамина D3). Кальцитриол оказывает воздействие на тонкий кишечник, почки и кости. В клетках кишечника кальцитриол индуцирует синтез Са2+-переносящих белков — кальбайндинов, которые обеспечивают всасывание ионов кальция и фосфатов из полости кишечника в кровь, благодаря чему концентрация ионов кальция во внеклеточной жидкости поддерживается на уровне, необходимом для минерализации органического матрикса костной ткани. В почках кальцитриол стимулирует реабсорбцию ионов кальция и фосфатов. Эффекты кальцитриола на костную ткань связаны с прямой стимуляцией остеобластов и обеспечением костной ткани усиленно всасывающимся в кишечнике кальцием, что активирует рост и минерализацию кости. Гормоны коры надпочечников: глюкокортикоиды и минералокортикоиды. Химическая природа. Биологическая роль. Надпочечники Состоит из двух частей: 1) Коркового слоя – 90% массы надпочечников. 2) Мозгового – находится в центре, в окружении коры надпочечников. Кора состоит из 3 слоёв : 1) Клубочковый 2) Пучковый 3) Сетчатый В клубочковом слое синтезируются минералокортикоиды( основной представитель – альдестерон. В пучковом и сетчатом синтезируются глюкокортикоиды ( главный представитель – кортизон и синтезируются половые гормоны). Гормоны коры надпочечников являются стероидными , их синтез начинается с холестерола, который поступает в кору из крови, в составе липопротеинов низкой плотности или может синтезироваться в коре из ацетил-КоА. Функции коры надпочечников, особенно сетчатой и пусковой, зависит от АКТГ(аденокортикотропный гормон) в передней доле гипофиза. Кортизол секретируется в кровь в свободном состоянии, но сразу связывается с кортизолом, связывается с глобулином ( транскортин – белок переносит основную массу кортизола, также транспорт кортизола могут осуществлять альбумины) Функции кортизола Направлен на поддержание метаболизма липидов, углеродов и белков, на поддержание работы нервно-сосудистой системы, почек, деятельности ЦНС, иммунной системы, кортизол обеспечивает устойчивость организма к стрессу. Кортизол увеличивает скорость механизма в жировой ткани, путём повышения активности Катехоламины Увеличивает липолитический эффект , увеличивает синтез холестерола и липопротеинов очень низкой плотности в печени. Кортизол обладает катаболитическим процессом на белковый обмен, усиливает распад белков до свободных аминокислот, стимулирует образование глюкозы из неуглеродного продукта, т.е глюконеогенез, снижает чувствительность тканей к инсулину, гормон обладает иммуносупрессивным действием , оказывает цитотоксическое действие на клетки иммунной системы, подавляет выработку антител. Гормон усиливает функцию миокарда, увеличивает сердечный выброс, тонус артериол. В физиологических концентрации увеличивает обратимое всасывание натрия в почках, увеличивает объем внеклеточной жидкости, активирует секреторные железы желудка, кишечника, при стрессе увеличивается содержание кортизола, уменьшает количество антител. Кортизол тормозит активность фибробласт, снижает количество коллагена и соединительной ткани (увеличивает морщинистость). Кортизол свободно проходит через гематоэнцефалический барьер , влияет на поведение, на половую функциональность, влияет на гонадотропную активность гипофиза и функции почечных желез, тормозит секрецию половых гормонов. В клубочковой зоне синтезируются минералокортикоиды, основным представителем является альфастерол – образуется из холестерола, кровью транспортируются с альбумином, образуется цитозольным механизмом действия. Гормон регулирует водно-солевой обмен, мишенью являются почки, гормон обеспечивает синтез белков и ферментов, котрые обеспечивают открытие калиевых и натриевых каналов, собирательных трубочек почек и движение в них натрия, это происходит при участии натрий-калиевая АТФ-фазаьи альдестерон регулирует кеназы, в результате концентрация натрия в кровь увеличивается, а калия уменьшается, в моче происходит наоборот. Регуляция водно-солевого обмена. Химическая природа и функции альдостерона и вазопрессина. Система ренин-ангиотензин и вазопрессин. Ангиотензин-превращающий фермент. Биохимические механизмы возникновения гипертонии, отеков, дегидратации. Регуляция водно-солевого обмена. Химическая природа и функции альдостерона и вазопрессина. Основными эффекторными регуляторами водно-солевого обмена являются альдостерон, натрийуретические пептиды и вазопрессин. Альдостерон Стероидный гормон альдостерон, самый активный минералокортикоид, секретируемый клубочковой зоной коры надпочечников. В клубочковой зоне коры надпочечников синтезируется минералкортикоиды, основным представителем является альдостерон, образованный из холлесторола. В крови транспортируется с альбумином. Обладает цитозольным механизмом действия. Гормон регулирует ВОДНО-СОЛЕВОЙ обмен. Мишенью являются почки. Гормон обеспечивает Синтез Белков и ферментов, которые регулируют открытие натрий калиевых каналов собирательных трубочек почек и движение в них натрия. Это происходит при участии натрий калиевой АТФ-фазы и АЛЬДОСТЕРОНРЕГУЛИРУЮЩЕЙ киназы. В результате концентрация Na в крови увеличивается,а K уменьшается(в моче наоборот). В почках альдостерон усиливает реабсорбцию Na+ в собирательных трубочках и в дистальных извитых канальцах (частично), усиливает выведение H+ и К+. На системном уровне альдостерон вызывает: Задержку Na+ и Cl-. Задержку воды. Выведение H+ и К+ почками. Увеличение ОЦК и АД. Поддержание водно-электролитного баланса. Вазопрессин (антидиуретический гормон/АДГ) В гипоталамусе. Регулирует ВОДНО-СОЛЕВОЙ обмен, мишенью для него являются сосуды и почки. Действуя на почки гормон повышает проницаемость собирательных трубочек для воды, действия на сосуды, он вызывает их сокращение конечным результатом действия гормона являются осмомолярность мочи, снижение её объема и сужение сосудов, также гормон действует на ЦНС, снижение температуры, облегчает консолидация памяти. Рецепторы вазопрессина – мембранные рецепторы, сопряжённые с G-белками. Функции Стимуляция реабсорбции воды: – Активация синтезированных аквапоринов. – Повышение экспрессии гена AQP2. Увеличение АД (вазоконстрикция + реабсорбция воды). Стимуляция секреции АКТГ аденогипофизом. Социальное поведение (слабее, чем окситоцин). Система ренин-ангиотензин и вазопрессин. РААС – физиологическая система, призванная поддерживать в организме оптимальное АД в покое и при кровопотере. Ключевой регулятор РААС – объём циркулирующей крови (ОЦК). Снижение ОЦК провоцирует увеличение секреции юкстагломерулярным аппаратом почки фермента ренина105. Попадая в кровь, ренин отщепляет от ангиотензиногена, секретируемого печенью, декапептид ангиотензин I. Под действием тканевого106 ангиотензинпревращающего фермента (АПФ) ангиотензин I превращается в октапептид ангиотензин II (ATII). ATII– физиологическое гипертензивное соединение, эффективно поднимающее АД благодаря вазоконстрикции и стимуляции синтеза альдостерона. Другой стимулятор секреции альдостерона – концентрация К+ в крови. Альдостерон регулирует свою продукцию по механизму отрицательной обратной связи, угнетая активность верхних звеньев РААС и снижая плазменную концентрацию К+. Он увеличивает задержку Na+, восстанавливая ОЦК, что, в свою очередь, угнетает продукцию ренина. Возросшая при гиперкалиемии продукция альдостерона ускоряет экскрецию К+. Затем, вслед за снижением концентрации К+ в плазме, падает и продукция альдостерона (Рис. 67). Секреция альдостерона также снижается под действием натрийуретических пептидов и дофамина. 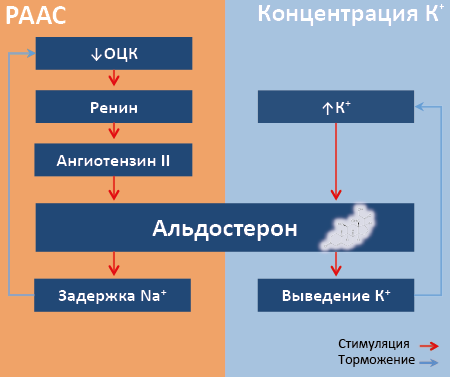 Ангиотензин-превращающий фермент Ангиотензин-превращающий фермент (АПФ) представляет собой мембраносвязанную пептидазу, содержащуюся в различных органах и тканях, в наибольшем количестве в клетках легких. Ангиотензин-превращающий фермент является ключевым ферментом ренин-ангиотензин-альдостероновой системы, где он расщепляет ангиотензин I с образованием активного вазоконстриктора ангиотензина IIАнгиотензин-превращающий фермент (АПФ) представляет собой мембраносвязанную пептидазу, содержащуюся в различных органах и тканях, в наибольшем количестве. АПФ может гидролизовать многие пептидные субстраты, обладая различной экзо- и эндопептидазной активностью. Основную физиологическую функцию АПФ в организме связывают с регуляцией кровяного давления, однако вероятно его участие и во множестве других физиологических и патологических процессов клетки в почках, мужской репродуктивной системе, иммунной системе. Биохимические механизмы возникновения гипертонии, отеков, дегидратации. Гиперальдостеронизм - заболевание, вызванное гиперсекрецией альдостерона надпочечниками. Причиной первичного гиперальдостеронизма (синдром Кона) примерно у 80% больных является аденома надпочечников, в остальных случаях - диффузная гипертрофия клеток клубочковой зоны, вырабатывающих альдостерон. При первичном гиперальдостеронизме избыток альдостерона усиливает реабсорбцию натрия в почечных канальцах. Увеличение концентрации Na+ в плазме служит стимулом к секреции АДГ и задержке воды почками. Кроме того, усиливается выведение ионов калия, магния и протонов. В результате развиваются гипернатриемия, вызывающая, в частности, гипертонию, гипер-волемию и отёки, а также гипокалиемия, ведущая к мышечной слабости, возникают дефицит магния и лёгкий метаболический алкалоз. Вторичный гиперальдостеронизм встречается гораздо чаще, чем первичный, и может быть связан с рядом состояний (например, сердечная недостаточность, хронические заболевания почек, а также сопровождающиеся нарушением кровоснабжения опухоли, секретирующие ренин). При вторичном гиперальдостеронизме у больных наблюдают повышенный уровень ренина и ангиотензина II, что стимулирует кору надпочечников продуцировать и секретировать избыточное количество альдостерона. Клинические симптомы менее выражены, чем при первичном альдостеронизе. Одновременное определение концентрации альдостерона и активности ренина в плазме позволяет окончательно дифференцировать первичный (активность ренина в плазме снижена) и вторичный (активность ренина в плазме повышена) гиперальдостеронизм. Роль предсердного натрийуретического фактора в регуляции осмотического давления и артериального давления. Предсердный натрий уретический фактор имеет белковую природу. Синтезируется предсердиями в ответ на растяжение. Действует на почки, увеличивает тонус приносящих артериол, тем самым повышает давление в клубочках. Способствует увеличению выделения Na, подавляет секрецию вазопрессина. Конечные эффекты гормона направлены на возврат к норме увеличенного количества Na и объёма воды в организме. Гормоны мозгового слоя надпочечников. Их химическая природа и биологическая роль. Мозговой слой надпочечников находится в центральной части надпочечников. В мозговом слое синтезируется катехоламины(КА), дофамины, норадреналин(НА) адреналин(АДР). Являются производными аминокислоты тирозина, обладающим мембранным механизмом действия. Основной гормон- адерналин. Нораденалин –нейромедиатор в симпатичекой нервной системе в постганглионарных волокнах. Рецепторы , с которыми связывается гормон – адренорецепторами ( альфа и бета ): Альфа адренорецепторы (сужение сосудов) – регулируют тонус сосудов, перистальтику кишечника, расширение зрачка. Бета адренорецеторы (альфа сужение, бета расширение) – влияют на силу и частоту сокращения миокарда ,тонус сосудов, тонус бронхов( расширение) и липолиз Катехоламины действуют на сердечно-сосудистую систему, поддерживая нужный кровоток коронарных артерий. Действуют через бета рецепторы на сердце, увеличивают силу и частоту сердечных сокращений, увеличивается потребность сердца в кислороде. Катехоламины в печени и мышцах стимулируют распад гликогена, в жировой ткани стимулирует липолиз, на внутренние органы действует через гладкую мускулатуру, вызывающее расширение бронхов, расслабление гладкой мускулатуры мочевого пузыря, кишечника. Гормоны поджелудочной железы и желудочно-кишечного тракта. Строение, синтез и секреция инсулина. Биологические функции и механизм действия инсулина. Строение и биологическая роль глюкагона. Поджелудочная железа- смешаннач секреция. Часть ее синтезирует пищеварительные ферменты, которые поступают в двенадцатиперстную кишку , а эндокринная часть представлена островками Лангерганца, которая состоит из клеток разоичного типа: •альфа клетки синтезируют гормон гликогон •бета клетки синтез инсулин •гамма клетки синтез соматостатийные Инсулин - гормон белковой природы , состоит из двух полипептидных цепей, соединенных дисульфидными связями Инсулин - анаболитический гормон, который обеспечивает транспорт глюкозы, аминокислот , ионов, клетки организма( особенно печени ), почек и дировой ткани. Инсулин обеспечивает утилизацию глюкозы в клетках путем увеличения активночти ферментов глюкокиназы и гексокиназы. В результате глюкоза фосфорилируется с образованием глюкози 6фосфата , которое может в зависимости от потребности организма окисляться , превращаться в жиры , либо накапливаться в виде гликогена. Общий результат этих процессов - снижение содержания глюкозы в крови. Инсулин -единственный гормон , снижаюший содержание глюкозы в крови. Глюкоза транспортируется прежде всего в печень, мышцах и жировую ткань. В печени и жировой ткани гормон стимулирует синтез жиров, тормозит их распад , в результате чего концентрация жирных кислот крови снижается. Инсулин стимулирует потребление аминокислот мышцами. При нарушении структуры и секреции развивается заболевание -сахарный диабет. Уменьшается депонирование гликогена, жиров; усиливается распад энергосистем для сахарного диабета: гипергликимия( повышение содержания глюкозы в крови) , глбкозория (глюкоза выделяется с мочой ), кетонемия (повешение концентрации кетоновых тел в крови, накопление кетоновых тел приводит к ацидозу). При сахарном диабете снижается скорость синтеза белков , ускоряется их распад. Это приводит к увел аминокислот в крови поступают в печень, там дезаминируется. Их безазотистый остаток включ в процесс глюконеогенеза ,что усиливает гипергликемию. При этом образовавшийся аммиак вступает в орнитиновый цикл с образованием мочевины, что приводит к увеличению ее в крови и моче. Высоко концентрированные глюкоза , кетоновые тела и мочевина требует выведения из организма. Но концентрированная способность почек ограничена , поэтому увел выделение большого количества воды. В результате может наступить обезвоживания организма, т.е при диабете наблюдается полиурия (выделение большого количества мочи ), а потеря воды вызывает постоянную жажды – полидиксию. Глюкогон - полипептид , он явл антаганистом инсулина. Повышает уровень глюкозы в крови, стимулируя распад гликогена, стимулирует глюкогенез. В клетках жировой ткани стимулируется липолиз (распад липидов ) , т.е противоположность липидов глюкогон стимулирующий мобилизацию осн энергоносителей , углеводов и жиров. В дельта клетках стимулируется -соматостатин , который регулирует выведение инсулина и гликогона. Гормоны ЖКТ    Регуляция энергетического обмена. Изменения метаболизма в абсорбтивный и постабсорбтивный периоды. Изменения гормонального статуса и метаболизма при голодании. Центральную роль в этом играет гипоталамус который интегрирует афферентные сигналы и способствует возникновению эфферентных сигналов, регулирует пищевое поведение и энергозатраты. В регуляции энергетического обмена участвует инсулин, глюкогон и теревоидные гормоны. Изменения гормонального статуса и метаболизма при сахарном диабете. При нарушении структуры и секреции инсулина развивается заболевание сахарный диабет, уменьшение депонирования гликогена, жиров, усиливается распад энергоносителей . для сахарного диабета характерны гипергликемия (повышение глюкозы), глюкозурия (выделения глюкозы с мочой), кетономия (повышение концентрации кетоновых тел в крови), накопление кетоновых тел приводит к ацидозу (повышение кислотности). При сахарном диабете снижается скорость синтеза белков, ускоряется их распад. Это приводит к увеличению аминокислот в крови, аминокислоты поступают в печень, там дизламинируются. Их без азотистый остаток включается в процесс глюканалагенеза что ещё больше усиливает гипергликемию. При этом образуется аммиак вступает в орнетиновый цикл с образованием мочевины что приводит к её увеличению в крови и моче. Высокая концентрация глюкозы, кетоновых тел и мочевины требуют усиленного выведения из организма, но но концентрационность почек ограничена поэтому увеличивается выделение большого количества воды, в результате может наступить обезвоживание организма. То есть при диабете наблюдается полиурия (выделения большого количества мочи) а потея воды вызывает постоянную жажду полидипсию Структура и биологическая роль важнейших тканевых гормонов. Тканевые гормоны - это гормоны, которые обладают паро- и аудо-действием или те, которые открылись недавно и про них мало известно. Лептин- гормон жировой ткани. Открыт американскими учеными в 1994 году. Помимо адипоцитов, может синтезироваться слизистой оболочкой желудка. По химической природе полипептид. Кровью транспортируется в свободной форме и в связанной форме с альбуминами. Поступая в кровь, проникает через гематоэнцефалический барьер в мозг, где связывается с рецепторами гипоталамуса (проходит в мозг, потому что он маленький). Стимулирует чувства насыщения, снижает чувство голода и потребление пищи. Может взаимодействовать с холецистокинином жкт из способствовать формированию чувства насыщения. Гормон препятствует снижению энергетических запасов в организме, предотвращает развития ожирения, оказывают влияние на адипоциты, предотвращая липидогенез (синтез липидов), стимулирует липолиз (распад липидов) в мышцах, увеличивает окисления жирных кислот, таким образом формирует липотоксин. Адипонектин- открыт в 1995, 1996 годах. Синтез в подкожный жировой ткани. Печени увеличивать чувствительность к инсулину, глюконеогенез, окисления жирных кислот. В мышцах стимулирует окисления глюкозы и жирных кислот. Влияет на стенки сосудов, ингибируя адгезию моноцитов, трансформацию макрофага в пенные клетки, таким образом препятствует развитию атеросклероза. Стимулирует продукцию оксида азота и стимулирует ангиогенез (синтез капилляров). Висфатин- открыт в 2004 году японскими учеными. Синтезируется жировой тканью. Обладает свойствами цитокинов и свойствами фермента, участвующего в биосинтезе NAD, таким образом участвуют во многих биохимических процессах. Обладает иммуномодулирующим эффектом. Помимо жировой ткани, синтезируется макрофагами и при взаимодействии с сосудистыми стенками с прoвоспалительными агентами, активизирует лейкоциты и стимулирует синтез цитокинов. Грелин- открыт в 1999 году японскими учеными. Является пептидом. Клетки продуцирующие гормон находится в желудке, в легких, в почках, в поджелудочной железе и в кишечнике. Гормон синтезируется и в гипоталамусе, влияя на гипофиз и стимулируя синтез гормона роста. Связан с повышением аппетита, перед приемом пищи его концентрация высокая, после - низкая. Усиливает пищевое поведение, увеличивать частоту приемов пищи, влияет на работу гипокампа, активизирует синтез оксида азота, тормозит секрецию инсулина. Ангиотензин- является пептидом. Образуется из ангиотензиногена ( белок, синтезируемые печенью). Ангиотензиноген под влиянием ренина превращается в ангиотензин 1. Ренин синтезируется почками в ответ на снижение внутрипочечного давления и снижение доставки натрия и хлора. Ангиотензин 1 биоактивностью не обладает. Под влиянием ангиотензинпревращающего фермента (АПФ) превращается в биоактивной ангиотензин 2. Ангиотензин обладает сильным сосудосуживающим эффектом, повышает артериальное давление, регулирует агрегацию и адгезию тромбоцитов, вызывает чувство жажды, увеличивает синтез вазопрессина и секрецию альгостерона (влияет на водно солевой обмен). Эритропоэтин- является гликопротеина. Образуется в почках и печени. Контролирует дифференцировку стволовых клеток костного мозга, способствует образованию эритроцитов, гемоглобина. Предсердный натрийуретический фактор- имеют белковую природу. Синтезируется предсердиями в ответ на растяжение. Действует на почки, увеличивает тонус ... артериол, таким образом увеличивает давление в клубочках. Способствует увеличению выделения натрия, подавляет секрецию вазопрессина. В целом, конечно эффект гормона направлен на возврат к норме увеличенного количества натрия и объема воды в организме. Мозговой натрийуретический фактор- не изучен, писать не будем))))) Эндотелин- является пептидом. Секретируется эндотелием. Является 1 из самых сильных веществ, с у живущих сосуды. Также стимулирует все этапы гемостаз (остановка кровотечения), агрегацию тромбоцитов. Гормоны, регулирующие кальций-фосфорный обмен Кальцитонин уменьшает концентрацию Са2+ в крови, способствует транспорту Са и Р из крови в костную ткань, подавляет реабсорбцию ионов Са2+ в почках, тем самым стимулируя экскрецию их с мочой. Кальцитонин по характеру действия является антагонистом паратгормона. Он ингибирует высвобождение Са2+ из кости, снижая активность остеокластов. Паратгормон – основное действие паратгормона направлено на повышение концентрации ионов кальция и снижение концентрации фосфатов в плазме крови. Органы-мишени для него – кости и почки. В костной ткани паратгормон ускоряет образование ферментов щелочной фосфатазы и коллагеназы, которые воздействуют на компоненты костного матрикса, вызывают его распад, в результате чего происходит мобилизация Са2+ и фосфатов из кости в кровь. В почках паратгормон стимулирует реабсорбцию кальция в извитых канальцах и тем самым задерживает его в организме, уменьшая экскрецию его с мочой, уменьшает реабсорбцию фосфатов. Кальцитриол оказывает воздействие на тонкий кишечник, почки и кости. В клетках кишечника кальцитриол индуцирует синтез Са2+-переносящих белков — кальбайндинов, которые обеспечивают всасывание ионов кальция и фосфатов из полости кишечника в кровь, благодаря чему концентрация ионов кальция во внеклеточной жидкости поддерживается на уровне, необходимом для минерализации органического матрикса костной ткани. В почках кальцитриол стимулирует реабсорбцию ионов кальция и фосфатов. Эффекты кальцитриола на костную ткань связаны с прямой стимуляцией остеобластов и обеспечением костной ткани усиленно всасывающимся в кишечнике кальцием, что активирует рост и минерализацию кости. |