Лекция 2 _Основы микробиологии_1 леч.дело. Лекция Физиология и биохимия микроорганизмов
 Скачать 241.54 Kb. Скачать 241.54 Kb.
|

|
Лекция № 2. Физиология и биохимия микроорганизмов Физиология микроорганизмов изучает жизнедеятельность микробных клеток, процессы их питания, дыхания, роста, размножения, закономерности взаимодействия с окружающей средой. Химический состав прокариотной клетки Клетки прокариот содержат воду, белки, нуклеиновые кислоты, углеводы, липиды, минеральные вещества . Вода — основной компонент бактериальной клетки, составляющий около 70 - 90 % ее массы. Она находится в свободном или связанном состоянии со структурными элементами клетки. В спорах количество воды уменьшается до 18 - 20 %. Вода является растворителем для многих веществ, а также выполняет механическую роль в обеспечении тургора. При плазмолизе - потере клеткой воды в гипертоническом растворе - происходит отслоение протоплазмы от клеточной оболочки. Удаление воды из клетки, высушивание приостанавливают процессы метаболизма. Большинство микроорганизмов хорошо переносят высушивание. При недостатке воды они не размножаются. Высушивание в вакууме из замороженного состояния (лиофилизация) прекращает размножение и способствует длительному сохранению микробных особей. Остальные компоненты составляют 10 - 30 % сухой массы клетки. Белки определяют важнейшие биологические свойства бактерий и состоят обычно из сочетаний 20 аминокислот. В состав бактерий входит диаминопимелиновая кислота (ДАП), отсутствующая в клетках человека и животных. Бактерии содержат более 2000 различных белков, находящихся в структурных компонентах и участвующих в процессах метаболизма. Большая часть белков обладает ферментативной активностью. Белки бактериальной клетки обусловливают антигенность и иммуногенность, вирулентность, видовую принадлежность бактерий. 59 Нуклеиновые кислоты бактерий выполняют функции, аналогичные функциям нуклеиновых кислот эукариотических клеток: молекула ДНК в виде хромосомы отвечает за наследственность, рибонуклеиновые кислоты (информационная, или матричная, транспортная и рибосомная) участвуют в биосинтезе белка. Бактерии можно характеризовать (таксономически) по содержанию суммы гуанина и цитозина (ГЦ) в молярных процентах (мол. %) от общего количества оснований ДНК. Более точной характеристикой микроорганизмов является гибридизация их ДНК. Основа метода гибридизации ДНК - способность денатурированной (однонитчатой) ДНК ренатурироваться, то есть соединяться с комплементарной нитью ДНК и образовывать двухцепочечную молекулу ДНК. 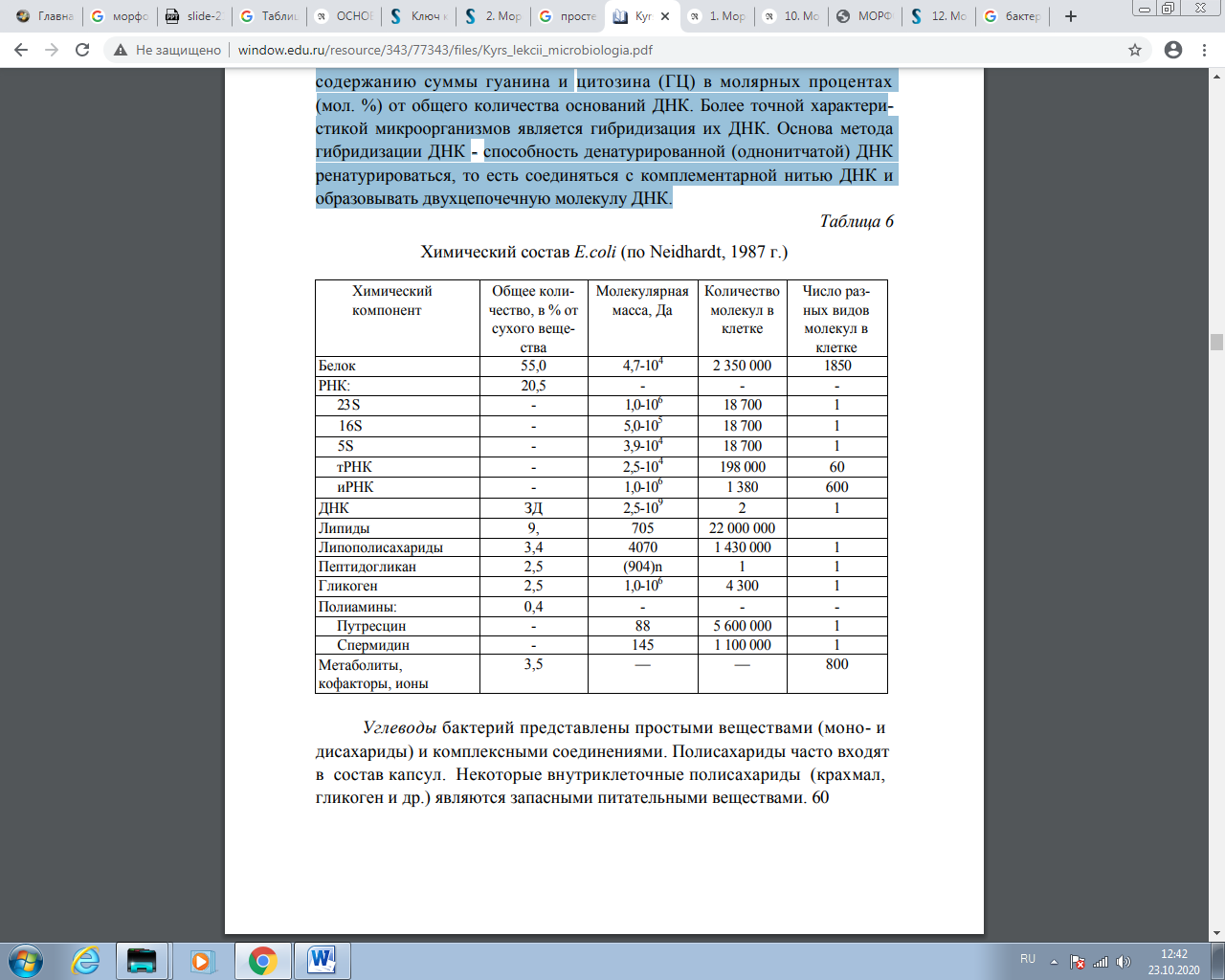 Углеводы бактерий представлены простыми веществами (моно- и дисахариды) и комплексными соединениями. Полисахариды часто входят в состав капсул. Некоторые внутриклеточные полисахариды (крахмал, гликоген и др.) являются запасными питательными веществами. Липиды в основном входят в состав цитоплазматической мембраны и ее производных, а также клеточной стенки бактерий, например наружной мембраны, где, кроме бимолекулярного слоя липидов, имеется ЛПС. Липиды могут выполнять в цитоплазме роль запасных питательных веществ. Липиды бактерий представлены фосфолипидами, жирными кислотами и глицеридами. Наибольшее количество липидов (до 40 %) содержат микобактерии туберкулеза. Минеральные вещества бактерий обнаруживают в золе после сжигания клеток. В большом количестве выявляются фосфор, калий, натрий, сера, железо, кальций, магний, а также микроэлементы (цинк, медь, кобальт, барий, марганец и др.). Они участвуют в регуляции осмотического давления, рН среды, окислительно-восстановительного потенциала, активируют ферменты, входят в состав ферментов, витаминов и структурных компонентов микробной клетки. Питание бактерий (конструктивный метаболизм прокариот). Образ жизни прокариот состоит в постоянном воспроизведении своей биомассы. Совокупность протекающих в клетке процессов, обеспечивающих воспроизводство биомассы, называется обменом веществ, или метаболизмом. Клеточный метаболизм складывается из двух потоков реакций, имеющих разную направленность: энергетического и конструктивного метаболизмов. КОНСТРУКТИВНЫЙ МЕТАБОЛИЗМ Конструктивный метаболизм (биосинтез, анаболизм) - цепь последовательных реакций, в результате которых за счет поступающих извне веществ строится вещество клеток; это процесс, связанный с потреблением свободной энергии, запасенной в химической форме в молекулах АТФ или других богатых энергией соединениях. Метаболические пути конструктивной и энергетической направленности состоят из множества последовательных ферментативных реакций и могут быть разделены на три этапа: 1) На начальном этапе воздействию подвергаются молекулы, служащие исходными субстратами. Иногда эту часть метаболического пути называют периферическим метаболизмом, а ферменты, катализирующие первые этапы превращения субстрата, - периферическими; 2) Последующие превращения включают ряд ферментативных реак ций и приводят к образованию промежуточных продуктов, или метаболи тов, а сама цепь превращений объединяется под названием промежуточно го метаболизма; 3) На последних этапах образуются конечные продукты конструк тивных путей, которые используются для построения вещества клеток, а конечные продукты энергетических путей выделяются в окружающую среду. Конструктивные и энергетические процессы протекают в клетке одновременно. У большинства прокариот они тесно связаны между собой. Однако у некоторых прокариотных организмов можно выделить последовательности реакций, служащих только для получения энергии или только для биосинтеза. Связь между конструктивными и энергетическими процессами прокариот осуществляется по нескольким каналам. Основной из них - энергетический. Определенные реакции поставляют энергию, необходимую для биосинтезов и других клеточных энергозависимых функций. Биосинтетические реакции, кроме энергии, нуждаются часто в поступлении извне восстановителя в виде водорода (электронов), источником которого служат также реакции энергетического метаболизма. И наконец, тесная связь между энергетическими и конструктивными процессами проявляется в том, что определенные промежуточные этапы и метаболиты обоих путей могут быть одинаковыми (хотя направленность потоков реакций, относящихся к каждому из путей, различна). Это создаѐт возможности для использования общих промежуточных продуктов в каждом из метаболических путей. Промежуточные соединения такой природы предложено называть амфиболитами, а промежуточные реакции, одинаковые для обоих потоков, - амфиболическими. Метаболизм прокариот отличается чрезвычайным разнообразием, которое есть результат способности этих форм жизни использовать в качестве источников энергии и исходных субстратов для построения веществ тела самый широкий набор органических и неорганических соединений. Такая способность обусловлена различиями в наборе клеточных периферических ферментов, воздействующих на исходные субстраты и видоизменяющих их молекулы в направлении, позволяющем им далее метаболизироваться по каналам промежуточного метаболизма. Промежуточный метаболизм прокариот не отличается существенным разнообразием от пери ферического метаболизма эукариот, хотя по сравнению с ним состоит из большего числа вариантов. Мономеры, необходимые для построения основных клеточных компонентов, могут быть синтезированы клеткой или поступать в готовом виде из среды. Чем больше готовых соединений должен получать организм извне, тем ниже уровень его биосинтетических способностей, так как химическая организация всех свободноживущих форм одинакова. ПИТАНИЕ БАКТЕРИЙ Особенности питания бактериальной клетки состоят в поступлении питательных субстратов внутрь через всю ее поверхность, а также в высокой скорости процессов метаболизма и адаптации к меняющимся условиям окружающей среды. Широкому распространению бактерий способствует разнообразие типов питания. Микроорганизмы нуждаются в углероде, азоте, сере, фосфоре, калии и других элементах. Источники углерода. В конструктивном метаболизме основная роль принадлежит углероду, поскольку все соединения, из которых построены живые организмы, - это соединения углерода. Их известно около миллиона. Прокариоты способны воздействовать на любое известное углеродное соединение, то есть использовать его в своем метаболизме. В зависимости от источника углерода для конструктивного метаболизма все прокариоты делятся на две группы: автотрофы, к которым принадлежат организмы, способные синтезировать все компоненты клетки из углекислоты (аутотрофными бактериями являются нитрифицирующие бактерии, находящиеся в почве; серобактерии, обитающие в воде с сероводородом; железобактерии, живущие в воде с закисным железом, и др.) и гетеротрофы, для которых источником углерода служат органические соединения. Понятия «авто-» и «гетеротрофия» характеризуют, таким образом, тип конструктивного метаболизма. Если автотрофия - довольно четкое и узкое понятие, то гетеротрофия - понятие весьма широкое и объединяет организмы, резко различающиеся своими потребностями в питательных веществах. Наибольшая степень гетеротрофности присуща прокариотам, относящимся к облигатным внутриклеточным паразитам (от греч. parasitos нахлебник), то есть организмам, которые могут жить только внутри других живых клеток, например, риккетсии, вирусы и некоторые простейшие. Они являются патогенными для растений, животных и человека. Паразитический образ жизни привел к редукции некоторых метаболических путей у этих прокариот, что и обусловило полную их зависимость от метаболизма клетки хозяина. Факультативными паразитами (или условно-патогенными) называют формы прокариот, способные расти при создании подходящих условий вне клетки хозяина. Следующую крупную группу прокариот составляют гетеротрофы, утилизирующие органические остатки отмерших организмов в окружающей среде, их называют сапрофитами. Термин «сапрофиты» происходит от греческих слов sapros - гнилой и phyton - растение. Сапрофиты используют продукты жизнедеятельности других организмов или разлагающиеся растительные и животные ткани. К сапрофитам относится большая часть бактерий. Степень требовательности к субстрату у сапрофитов весьма различна. В эту группу входят организмы, которые могут расти только на достаточно сложных субстратах (молоко, трупы животных, гниющие растительные остатки), то есть им нужны в качестве обязательных элементов питания углеводы, органические формы азота в виде набора аминокислот, пептидов, белков, все или часть витаминов, нуклеотиды или готовые компоненты, необходимые для синтеза последних (азотистые основания, пятиуглеродные сахара). Чтобы удовлетворить потребность этих гетеротрофов в элементах питания, их обычно культивируют на средах, содержащих мясные гидролизаты, автолизаты дрожжей, растительные экстракты, молочную сыворотку. Есть прокариоты, требующие для роста весьма ограниченное число готовых органических соединений в основном из числа витаминов и аминокислот, которые они не в состоянии синтезировать сами, и, наконец, гетеротрофы, нуждающиеся только в одном органическом источнике углерода. Им может быть какой-либо сахар, спирт, кислота или другое углеродсодержащее соединение. Так, бактерии из рода Pseudomonas способны использовать в качестве единственного источника углерода и энергии любое из 200 различных органических соединений, и есть бактерии, для которых таким источником может служить узкий круг довольно экзотических органических веществ. Например, Bacillus fastidiosus может использовать только мочевую кислоту и продукты ее деградации, а некоторые представители рода Clostridium растут только в среде, содержащей пури ны. Использовать другие органические субстраты для роста они не могут. Биосинтетические способности этих организмов развиты в такой степени, что они сами могут синтезировать все необходимые им углеродные соединения. Особую группу гетеротрофных прокариот, обитающих в водоемах, составляют олиготрофные бактерии, способные расти при низких концентрациях в среде органических веществ. Организмы, предпочитающие высокие концентрации питательных веществ, относят к копиотрофам. Термины происходят от греческих слов oligos - малый, сорiosus - изобилие и trорhе - пища Если у типичных копиотрофов оптимальные условия для роста создаются при содержании в среде питательных веществ в количестве примерно 10 г/л, то для олиготрофных организмов - в пределах 1 - 15 мг углерода/л. В средах с более высоким содержанием органических веществ такие бактерии, как правило, расти не могут и погибают. Различия между гетеротрофными прокариотами с высокими потребностями в готовых органических соединениях и теми, потребности которых минимальны и сводятся, как правило, к одному какому-нибудь органическому источнику углерода, заключаются, таким образом, в степени развития их биосинтетических способностей. Крайняя степень развития биосинтетических способностей - возможность строить все клеточные компоненты из углекислоты - присуща группе автотрофных прокариот. Как можно видеть из изложенного выше, в мире прокариот не существует резкой границы между авто- и гетеротрофными организмами, так же как нет ее в ряду одноуглеродных соединений (СО2, СО, НСООН, НСНО, СН3ОН, СН4), каждое из которых может служить источником углерода для определенной группы прокариот. Однако использование термина «автотрофия» удобно для обозначения конкретного типа конструктивного метаболизма, поскольку в процессе эволюции он оказался специфически связанным с определенными видами энергетических процессов, что привело к появлению у прокариот таких типов жизни, которые отсутствуют у более высокоорганизованных форм. В зависимости от окисляемого субстрата, называемого донором электронов или водорода, микроорганизмы делятся на две группы: литотрофы (от греч. lithos - камень), использующие в качестве доноров водорода неорганические соединения, и органотрофы, использующие органические соединения. Потребности в источниках серы и фосфора. Сера входит в состав аминокислот (цистеин, метионин), витаминов и кофакторов (биотин, липоевая кислота, кофермент А и др.), а фосфор - необходимый компонент нуклеиновых кислот, фосфолипидов, коферментов. В природе сера находится в форме неорганических солей, главным образом сульфатов, в виде молекулярной (элементной) серы или входит в состав органических соединений. Большинство прокариот для биосинтетических целей потребляют серу в форме сульфата, который при этом восстанавливается до уровня сульфида. Однако некоторые группы прокариот не способны к восстановлению сульфата и нуждаются в восстановленных соединениях серы. Основной формой фосфора в природе являются фосфаты, которые и удовлетворяют потребности прокариот в этом элементе. Необходимость в ионах металлов. Всем прокариотным организмам необходимы металлы, которые могут использоваться в форме катионов неорганических солей. Некоторые из них (магний, кальций, калий, железо) нужны в достаточно высоких концентрациях, потребность в других (цинк, марганец, натрий, молибден, медь, ванадий, никель, кобальт) невелика. Роль перечисленных выше металлов определяется тем, что они входят в состав основных клеточных метаболитов и, таким образом, участвуют в осуществлении жизненно важных функций организма. Потребность в факторах роста. Некоторые прокариоты обнаруживают потребность в одном каком-либо органическом соединении из группы витаминов, аминокислот или азотистых оснований, которое они по каким-то причинам не могут синтезировать из используемого источника углерода. Такие органические соединения, необходимые в очень небольших количествах, получили название факторов роста. Факторы роста - это необходимые для микроорганизмов соединения, которые они сами синтезировать не могут, поэтому их необходимо добавлять в питательные среды. Организмы, которым в дополнение к основному источнику углерода необходим один или больше факторов роста, которые они не могут сами синтезировать, называют ауксотрофами (цв. вклейка, рис. IV, V, VI). Прототрофы способны сами синтезировать необходимые для роста соединения, например из глюкозы и солей аммония. МЕХАНИЗМЫ ПИТАНИЯ Поступление различных веществ в бактериальную клетку зависит от величины и растворимости их молекул в липидах или воде, рН среды, концентрации веществ, различных факторов проницаемости мембран и др. Клеточная стенка пропускает небольшие молекулы и ионы, задерживая макромолекулы массой более 600 Да. Основным регулятором поступления веществ в клетку является цитоплазматическая мембрана. Условно можно выделить четыре механизма проникновения питательных веществ в бактериальную клетку: простая диффузия, облегченная диффузия, активный транспорт, транслокация групп. Наиболее простой механизм поступления веществ в клетку - простая диффузия, при которой перемещение веществ происходит вследствие разницы их концентрации по обе стороны цитоплазматической мембраны. Вещества проходят через липидную часть цитоплазматической мембраны (органические молекулы, лекарственные препараты) и реже по заполненным водой каналам в цитоплазматической мембране. Пассивная диффузия осуществляется без затраты энергии. Облегченная диффузия происходит также в результате разницы концентрации веществ по обе стороны цитоплазматической мембраны. Однако этот процесс осуществляется с помощью молекул-переносчиков, локализующихся в цитоплазматической мембране и обладающих специфичностью. Каждый переносчик транспортирует через мембрану соответствующее вещество или передает другому компоненту цитоплазматической мембраны - собственно переносчику. Белками-переносчиками могут быть пермеазы, место синтеза которых - цитоплазматическая мембрана. Облегченная диффузия протекает без затраты энергии, вещества перемещаются из области с более высокой концентрацией в область с более низкой. Активный транспорт происходит с помощью пермеаз и направлен на перенос веществ меньшей концентрации в сторону большей, то есть как бы против течения, поэтому данный процесс сопровождается затратой метаболической энергии (АТФ), образующейся в результате окислительновосстановительных реакций в клетке. Перенос (транслокация) групп сходен с активным транспортом, но отличается тем, что переносимая молекула видоизменяется в процессе переноса, например фосфорилируется. Выход веществ из клетки осуществляется за счет диффузии и при участии транспортных систем. Дыхание прокариот (энергетический метаболизм) Энергетический метаболизм (катаболизм) - это поток реакций, сопровождающихся мобилизацией энергии и преобразованием ее в электрохимическую (А(Хн+) или химическую (АТФ) форму, которая затем может быть использована во всех энергозависимых процессах. Существуют группы прокариот, энергетический метаболизм которых не связан с превращениями органических соединений (прокариоты с фотолито- и хемолитотрофным типом энергетического обмена). По отношению к такого рода энергетическим процессам термин «катаболизм» неприменим. У них функционирует только один поток превращений органических соединений углерода - анаболический. Энергетические процессы прокариот по своему объему (масштабности) значительно превосходят процессы биосинтетические, а их протекание приводит к существенным изменениям в окружающей среде. Разнообразны и необычны в этом отношении возможности прокариот, способы их энергетического существования. Все это вместе взятое сосредоточило внимание исследователей в первую очередь на изучении энергетического метаболизма прокариот. Дыхание бактерий. Дыхание, или биологическое окисление, основано на окислительно-восстановительных реакциях, идущих с образованием АТФ-универсального аккумулятора химической энергии. Энергия необходима микробной клетке для ее жизнедеятельности. При дыхании происходят процессы окисления и восстановления: окисление - отдача донорами (молекулами или атомами) водорода или электронов; восстановление - присоединение водорода или электронов к акцептору. Акцептором водорода или электронов может быть молекулярный кислород (такое дыхание называется аэробным) или нитрат, сульфат, фумарат (такое дыхание называется анаэробным - нитратным, сульфатным, фумаратным). Анаэробиоз (от греч. aer - воздух, bios - жизнь) - жизнедеятельность, протекающая при отсутствии свободного кислорода. Если донорами и акцепторами водорода являются органические соединения, то такой процесс называется брожением. При брожении происходит ферментативное расщепление органических соединений, преимущественно углеводов, в анаэробных условиях. С учетом конечного продукта расщепления углеводов различают спиртовое, молочнокислое, уксусное и другие виды брожения. По отношению к молекулярному кислороду бактерии можно разделить на три основные группы: облигатные, то есть обязательные, аэробы, облигатные анаэробы и факультативные анаэробы. Облигатные аэробы - это прокариоты, для роста которых кислород необходим. К ним относится большинство прокариотных организмов. Облигатные анаэробы (клостридии ботулизма, газовой гангрены, столбняка, бактероиды и др.) растут только на среде без кислорода, который для них токсичен. При наличии кислорода бактерии образуют перекисные радикалы кислорода, в том числе перекись водорода и супероксиданион кислорода, токсичные для облигатных анаэробных бактерий, поскольку они не образуют соответствующие инактивирующие ферменты. Аэробные бактерии инактивируют перекись водорода и супероксид-анион кислорода соответствующими ферментами (каталазой, пероксидазой и супероксиддисмутазой). Факультативные анаэробы могут расти как при наличии, так и при отсутствии кислорода, поскольку они способны переключаться с дыхания в присутствии молекулярного кислорода на брожение в его отсутствие. Они способны осуществлять анаэробное дыхание, называемое нитратным: нитрат, являющийся акцептором водорода, восстанавливается до молекулярного азота и аммиака. Среди облигатных анаэробов различают аэротолерантные бактерии, которые сохраняются при наличии молекулярного кислорода, но не используют его. Для выращивания анаэробов в бактериологических лабораториях применяют анаэростаты - специальные емкости, в которых воздух заменяется смесью газов, не содержащих кислорода. Воздух можно удалять из питательных сред путем кипячения, с помощью химических адсорбентов кислорода, помещаемых в анаэростаты или другие емкости с посевами. Размножение вирусов ВЗАИМОДЕЙСТВИЕ ВИРУСА С КЛЕТКОЙ Известны три типа взаимодействия вируса с клеткой: 1) продуктивный тип, завершающийся образованием вирусного по томства; 2) абортивный тип, не завершающийся образованием новых вирус ных частиц, поскольку инфекционный процесс прерывается на одном из этапов; 3) интегративный тип, или вирогения, характеризующийся встраи ванием вирусной ДНК в хромосому клетки-хозяина. Продуктивный тип взаимодействия (репродукция вирусов). Репродукция вирусов (от англ. reproduce - воспроизводить) осуществляется в несколько стадий, последовательно сменяющих друг друга: - адсорбция вируса на клетке; - проникновение вируса в клетку; - «раздевание» вируса; биосинтез вирусных компонентов в клетке; - формирование вирусов; - выход вирусов из клетки. Адсорбция. Взаимодействие вируса с клеткой начинается с процесса адсорбции, то есть прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны - так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Количество специфических рецепторов на поверхности одной клетки колеблется от 104 до 105 . Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц. Поверхностные структуры вируса, «узнающие» специфические клеточные рецепторы и взаимодействующие с ними, называются прикрепительными белками. Обычно эту функцию выполняет один из поверхностных белков капсида или суперкапсида. Соответствие (комплементарность) клеточных рецепторов вирусным прикрепительным белкам имеет значение для возникновения инфекционного процесса в клетке. Способность вирусов избирательно поражать определенные клетки органов и тканей организма называют тропизмом вирусов (от греч. tropos - направление). ПРОНИКНОВЕНИЕ ВИРУСА В КЛЕТКУ. Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга. «Раздевание» вируса. Процесс «раздевания» заключается в удалении защитных вирусных оболочек и освобождении внутреннего компонента вируса, способного вызвать инфекционный процесс. «Раздевание» вирусов происходит постепенно, в несколько этапов, в определенных участках цитоплазмы или ядра клетки, для чего клетка использует набор специальных ферментов. В случае проникновения вируса путем слияния вирусной оболочки с клеточной мембраной процесс проникновения вируса в клетку сочетается с первым этапом его «раздевания». Конечными продуктами «раздевания» являются нуклеокапсид или нуклеиновая кислота вируса. Формирование (сборка) вирусов. Синтезированные вирусные нуклеиновые кислоты и белки обладают способностью специфически «узнавать» друг друга и при достаточной их концентрации самопроизвольно соединяются в результате гидрофобных, солевых и водородных связей. Существуют следующие общие принципы сборки вирусов, имеющих разную структуру: - формирование вирусов является многоступенчатым процессом с образованием промежуточных форм; - сборка просто устроенных вирусов заключается во взаимодейст вии молекул вирусных нуклеиновых кислот с капсидными белками и обра зовании нуклеокапсидов (например, вирусы полиомиелита). У сложно уст роенных вирусов сначала формируются нуклеокапсиды, с которыми взаи модействуют белки суперкапсидных оболочек (например, вирусы гриппа); - формирование вирусов происходит не во внутриклеточной жидко сти, а на ядерных или цитоплазматических мембранах клетки; - сложно организованные вирусы в процессе формирования вклю чают в свой состав компоненты клетки-хозяина (липиды, углеводы). Выход вирусов из клетки. Различают два основных типа выхода вирусного потомства из клетки. Первый тип - взрывной - характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имею щих суперкапсидной оболочки. Второй тип - почкование. Он присущ вирусам, имеющим суперкапсидную оболочку. На заключительном этапе сборки нуклеокапсиды сложно устроенных вирусов фиксируются на клеточной плазматической мембране, модифицированной вирусными белками, и постепенно выпячивают ее. В результате выпячивания образуется «почка», содержащая нуклеокапсид. Затем «почка» отделяется от клетки. Таким образом, внешняя оболочка этих вирусов формируется в процессе их выхода из клетки. При таком механизме клетка может продолжительное время продуцировать вирус, сохраняя в той или иной мере свои основные функции. Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5 - 6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инициировать новые клетки и проходить в них указанный выше цикл репродукции. |