|
|
7. Мочевыделительная система. Мужская и женская половая система.. Лекция Мочевыделительная система. Мужская и женская половая система к органам мочевыделительной системы относятся почки, мочеточники, мочевой пузырь и мочеиспускательный канал.
Лекция 7. Мочевыделительная система. Мужская и женская половая система
К органам мочевыделительной системы относятся почки, мочеточники, мочевой пузырь и мочеиспускательный канал. Среди них почки являются мочеобразующими органами, а остальные составляют мочевыводящие пути.
Развитие
В течение эмбрионального периода закладываются последовательно три парных выделительных органа:
• передняя почка (предпочка, pronephros);
• первичная почка (mesonephros);
• постоянная почка (окончательная, metanephros).
Предпочка образуется из передних 8-10 сегментных ножек (нефротомов) мезодермы.
У зародыша человека предпочка не функционирует в качестве мочеобразующего органа и вскоре после закладки подвергается атрофии.
Первичная почка (мезонефрос) формируется из большого числа сегментных ножек (около 25), расположенных в области туловища зародыша. Сегментные ножки, или нефротомы, отшнуровываются от сомитов и спланхнотома и превращаются в канальцы первичной почки. Канальцы растут по направлению к мезонефральному протоку, образующемуся еще при развитии предпочки, и вступают с ним в сообщение. Навстречу им от аорты отходят сосуды, распадающиеся на капиллярные клубочки. Канальцы своим слепым концом обрастают эти клубочки, образуя их капсулы. Капиллярные клубочки и капсулы вместе формируют почечные тельца. Возникший еще при развитии предпочки мезонефральный проток открывается в заднюю кишку.
Окончательная почка (метанефрос) закладывается у зародыша на 2-м месяце, но развитие ее заканчивается лишь после рождения ребенка. Эта почка образуется из двух источников — мезонефрального (Вольфова) протока и нефрогенной ткани, представляющей собой не разделенные на сегментные ножки участки мезодермы в каудальной части зародыша. Мезонефральный проток дает начало мочеточнику, почечной лоханке, почечным чашкам, сосочковым каналам и собирательным трубкам. Из нефрогенной ткани дифференцируются почечные канальцы. На одном их конце образуются капсулы, охватывающие сосудистые клубочки; другим концом они соединяются с собирательными трубками. Образовавшись, окончательная почка начинает быстро расти и с 3-го месяца оказывается лежащей выше первичной почки, которая во второй половине беременности атрофируется. С этих пор окончательная почка берет на себя все функции мочеобразования в организме плода.
ПОЧКИ
Почка (ren) — это парный орган, в котором непрерывно образуется моча. Почки регулируют водно-солевой обмен между кровью и тканями, поддерживают кислотно-щелочное равновесие в организме, а также выполняют эндокринные функции (включая регуляцию артериального давления и регуляцию эритропоэза).
Строение
Почка покрыта соединительнотканной капсулой и, кроме того, спереди — серозной оболочкой. Вещество почки подразделяется на корковое и мозговое. Корковое вещество (cortex renis) образует сплошной слой под капсулой органа. В процессе развития почки ее корковое вещество, увеличиваясь в массе, проникает между основаниями пирамид в виде почечных колонок (столбы Бертена). Мозговое вещество (medulla renis) состоит из 10-18 конических мозговых пирамид, от основания которых в корковое вещество врастают мозговые лучи.
Пирамида с покрывающим ее участком коры образует почечную долю, а мозговой луч с окружающим его корковым веществом - почечную дольку.
Строму почки составляет рыхлая волокнистая соединительная ткань (интерстиций). Паренхима почки представлена почечными тельцами и эпителиальными канальцами, которые при участии кровеносных сосудов образуют нефроны. В каждой почке их насчитывают около 1 млн.
Нефрон (nephronum) – это структурно-функциональная единица почки. Общая длина его канальцев достигает 5 см, а всех нефронов - около 100 км. Нефрон переходит в собирательную трубочку, которая продолжается в сосочковый канал, открывающийся на вершине пирамиды в полость почечной чашки.
Каждый нефрон включает: двустенную чашеобразную капсулу — капсулу Шумлянского-Боумена и отходящий от неѐ длинный эпителиальный каналец (с различными отделами). Концом нефрона считается место его впадения в одну изсобирательных почечных трубочек. Капсула Шумлянского-Боумена почти со всех сторон окружает капиллярный клубочек (glomerulus). Соответственно, почечное тельце (тельце Мальпиги) включает капиллярный клубочек и окружающую его капсулу.
От капсулы клубочка отходит проксимальный извитой каналец, делающий несколько петель возле почечного тельца. Проксимальный извитой каналец продолжается в петлю нефрона (петлю Генле). Нисходящая часть петли Генле (тонкий каналец) спускается вниз - по направлению к мозговому веществу (чаще всего, входя в него); восходящая часть (дистальный прямой каналец), более широкая, вновь поднимается по направлению к почечному тельцу нефрона.
В районе почечного тельца петля Генле переходит в дистальный извитой каналец. Дистальный извитой каналец одной своей петлѐй обязательно касается почечного тельца — между 2 сосудами (входящим и выходящим из клубочка на его вершине). Дистальный извитой каналец - последний отдел нефрона. Он впадает в собирательную почечную трубочку. Собирательные трубочки расположены почти перпендикулярно поверхности почки: вначале идут в составе мозговых лучей в корковом веществе, затем входят в мозговое вещество и у вершин пирамид впадают в сосочковые каналы, которые далее открываются в почечные чашки.
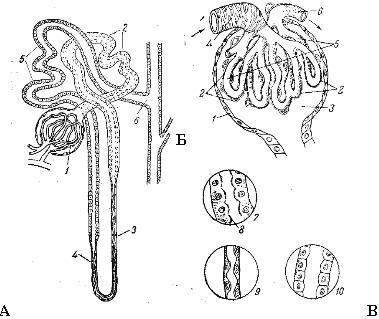
Схема строения нефрона (А), мальпигиева тельца(Б) и эпителия различных отделов канальца нефрона (В).
А: 1 - мальпигиево тельце; 2 - извитой каналец первого порядка (проксимальный); 3 - нисходящий отдел петли Генле; 4 - восходящий отдел петли Генле; 5 - извитой каналец второго порядка (дистальный); 6 - вставочный отдел нефрона; 7 - общая собирательная трубка.
Б: 1,2 - наружная и внутренняя стенки капсулы Шумлянского-Боумена; 3 - полость внутри капсулы; 4 - приносящая артериола; 5 - капиллярный клубочек; 6 - выносящая артериола; 7 - кубический эпителий проксимального канальца; 8 - микрореснички эпителиальных клеток; 9 - плоский эпителий петли Генле; 10 - эпителий дистального канальца.
Все почечные тельца лежат в корковом веществе. Извитые канальцы (проксимальный и дистальный) тоже находятся в коре, но положение петли Генле нефронов может существенно различаться. В связи с этим нефроны подразделяют на 3 типа:
1. Короткие корковые нефроны. Составляют не более1%от всех нефронов.Имеюточень короткую петлю, не достигающую мозгового вещества. Поэтому нефрон целиком лежит в коре.
2. Промежуточные корковые нефроны. Преобладают по численности( 80%всехнефронов). Часть петли «спускается» в наружную зону мозгового вещества.
3. Длинные (юкстамедуллярные, околомозговые) нефроны. Составляют не более20% всех нефронов. Почечные тельца их находятся в корковом веществе на границе с мозговым веществом. Петля Генле - очень длинная и почти целиком находится в мозговом веществе.
Таким образом, корковое и мозговое вещества почек образованы различными отделами трех разновидностей нефронов. Их топография в почках имеет определяющее значение для процессов мочеобразования, что в большой степени связано с особенностями кровоснабжения. В связи с наличием указанных типов нефронов в почке различают две системы кровообращения - кортикальную и юкстамедуллярную. Они совпадают в области крупных сосудов, но различаются ходом мелких сосудов.
Васкуляризация
Кровь поступает к почкам по почечным артериям, которые, войдя в почки, распадаются на междолевые артерии, идущие между мозговыми пирамидами. На границе между корковым и мозговым веществом они разветвляются на дуговые (аркуатные) артерии. От них в корковое вещество отходят междольковые артерии, от которых в стороны расходятся внутридольковые артерии. От этих артерий начинаются приносящие артериолы клубочков, причем от верхних внутридольковых артерий приносящие артериолы направляются к коротким и промежуточным нефронам (кортикальная система), от нижних - к юкстамедуллярным нефронам (юкстамедуллярная система).
Схема кровотока в кортикальной системе
Приносящая артериола входит в почечное тельце и распадается на 45-50 капиллярных петель (сосудистый клубочек, glomerulus), которые «распластываются» вблизи внутреннего листка капсулы и взаимодействуют с его клетками (см. ниже). Сформировав своими петлями «первичную» сеть, капилляры собираются в выносящую артериолу, которая покидает почечное тельце вплотную к месту вхождения приносящей артериолы (сосудистый полюс почечного тельца). Итак, на "входе" и на "выходе" из клубочка имеются две артериолы - приносящая (vas afferens) и выносящая (vas efferens), в результате чего «первичную» капиллярную сеть можно отнести к разряду rete mirabile (чудесных сетей). Важно подчеркнуть, что внутренний диаметр выносящей артериолы значительно уже, чем приносящей; благодаря этому создается своеобразный гемодинамический подпор крови в «первичной» сети и, как следствие, феноменально высокое давление крови в капиллярах — около 60 мм.рт.ст. Именно это высокое давление и является одним из главных условий основного процесса, происходящего в почечном тельце, - процесса фильтрации.
Выносящие артериолы, пройдя короткий путь, вновь распадаются на капилляры, оплетающие канальцы нефрона и образующие перитубулярную капиллярную сеть. В этих «вторичных» капиллярах давление крови значительно ниже, чем в «первичных» — около 10-12 мм.рт.ст., что способствует второй фазе мочеобразования - процессу реабсорбции (обратного всасывания) части жидкости и веществ из мочи в кровь. Из капилляров кровь перитубулярной сети собирается в верхних отделах коркового вещества сначала в звездчатые вены, а затем - в междольковые, в средних отделах коркового вещества - непосредственно в междольковые вены. Последние впадают в дуговые вены, переходящие в междолевые, которые образуют почечные вены, выходящие из ворот почек.
Таким образом, нефроны в связи с особенностями кортикального кровообращения (высокое давление крови в капиллярах сосудистых клубочков и наличие перитубулярной сети капилляров с низким давлением крови) активно участвуют в мочеобразовании.
Схема кровотока в юкстамедуллярной системе
Приносящие и выносящие артериолы сосудистых клубочков околомозговых нефронов примерно одинакового диаметра или выносящие артериолы даже несколько шире. Поэтому давление крови в капиллярах этих клубочков ниже, чем в клубочках корковых нефронов. Выносящие клубочковые артериолы юкстамедуллярных нефронов идут в мозговое вещество, распадаясь на пучки тонкостенных сосудов, несколько более крупных, чем обычные капилляры, - т.н. прямые сосуды (vasa recta). В мозговом веществе как от выносящих артериол, так и от прямых сосудов отходят ветви для формирования мозговой перитубулярной капиллярной сети. Прямые сосуды образуют петли на различных уровнях мозгового вещества, поворачивая обратно. Нисходящие и восходящие части этих петель образуют особую противоточную систему сосудов, называемую сосудистым пучком (fasciculus vasculans). Капилляры мозгового вещества собираются в прямые вены, впадающие в дуговые вены.
Вследствие этих особенностей околомозговые нефроны участвуют в мочеобразовании менее активно. В то же время юкстамедуллярное кровообращение играет роль шунта, т.е. более короткого и легкого пути, по которому проходит часть крови через почки в условиях сильного кровенаполнения, например, при выполнении человеком тяжелой физической работы.
Фильтрация
Фильтрация (главный процесс мочеобразования) происходит благодаря высокому давлению крови в капиллярах клубочков (50-60 мм.рт.ст.). В фильтрат (т.е первичную мочу) попадают многие компоненты плазмы крови - вода, неорганические ионы (например, Na+, K+, Cl- и другие ионы плазмы), низкомолекулярные органические вещества (в т.ч. глюкоза и продукты метаболизма - мочевина, мочевая кислота, желчные пигменты и др.), не очень крупные (до 50 кД) белки плазмы (альбумины, некоторые глобулины), составляющие 60-70 % всех плазменных белков. За сутки через почки проходит примерно 1800 л крови; из них в состав фильтрата перемещается почти 10 % жидкости. В итоге, суточный объѐм первичной мочи - около 180 л. Это более чем в 100 раз больше суточного объѐма конечной мочи (около 1,5 л). Следовательно, более 99 % воды, а также вся глюкоза, все белки, почти все прочие компоненты (кроме конечных продуктов обмена) должны возвращаться в кровь. Место, где разворачиваются все события процесса фильтрации — это почечное тельце.
Почечное тельце
Почечное тельце состоит из двух структурных компонентов - сосудистого клубочка и капсулы. Диаметр почечного тельца составляет в среднем 200 мкм. Сосудистый клубочек (glomerulus) состоит из 40-50 петель кровеносных капилляров. Их эндотелиальные клетки имеют многочисленные поры и фенестры (диаметром до 100 нм), которые занимают не менее 1/3 всей площади эндотелиальной выстилки капилляров. Эндотелиоциты располагаются на внутренней поверхности гломерулярной базальной мембраны. С наружной стороны на ней лежит эпителий внутреннего листка капсулы клубочка.
Капсула клубочка (capsula glomeruli) по форме напоминает двустенную чашу, образованную внутренним и наружным листками, между которыми расположена щелевидная полость — полость капсулы, переходящая в просвет проксимального канальца нефрона. Наружный листок капсулы — гладкий, внутренний - комплементарно повторяет контуры капиллярных петель, покрывая 80% площади поверхности капилляров. Внутренний листок образован крупными (до 30 мкм) неправильной формы эпителиальными клетками — подоцитами (podocyti — буквально: клетки с ногами, см. ниже).
Гломерулярная базальная мембрана, являющаяся общей для эндотелия кровеносных капилляров и подоцитов (и сформировавшаяся путем слияния эндотелиальной и эпителиальной базальных мембран), включает 3 слоя (пластинки): менее плотные (светлые) наружную и внутреннюю пластинки (laminae rara externa et interna) и более плотную (темную) промежуточную пластинку (lamina densa). Структурная основа темной пластинки представлена коллагеном IV типа, волокна которого формируют прочную решетку с размерами ячеек до 7 нм. Благодаря данной решетке темная пластинка играет роль механического сита, задерживающего частицы с большим диаметром. Светлые пластинки обогащены сульфатированными протеогликанами, которые поддерживают высокую гидрофильность мембраны и формируют ее отрицательный заряд, нарастающий и концентрирующийся от эндотелия и ее внутреннего слоя к наружному и к подоцитам. Данный заряд обеспечивает электрохимическое удерживание низкомолекулярных веществ, прошедших через эндотелиальный барьер. Помимо протеогликанов, светлые пластинки базальной мембраны содержат белок ламинин, обеспечивающий адгезию (прикрепление) к мембране ножек подоцитов и эндотелиоцитов капилляров.
Подоциты - клетки внутреннего листка капсулы - имеют характерную отросчатую форму: от центральной ядросодержащей части (тела) отходят несколько больших широких отростков 1-го порядка — цитотрабекул, от которых в свою очередь начинаются многочисленные мелкие отростки 2-го порядка - цитоподии, прикрепляющиеся к гломерулярной базальной мембране несколько утолщенными «подошвами» с помощью ламинина. Между цитоподиями располагаются узкие фильтрационные щели, сообщающиеся через промежутки между телами подоцитов с полостью капсулы. Фильтрационные щели шириной до 40 нм закрыты фильтрационными щелевыми диафрагмами. Каждая такая диафрагма — сеточка переплетающихся тончайших нитей из белка нефрина (ширина ячеек — от 4 нм до 7 нм), представляющая собой барьер для большинства альбуминов и других крупномолекулярных веществ. Кроме того, на поверхности подоцитов и их ножек имеется отрицательно заряженный слой гликокаликса, «усиливающий» отрицательный заряд базальной мембраны. Подоциты синтезируют компоненты гломерулярной базальной мембраны, образуют вещества, регулирующие кровоток в капиллярах и ингибирующие пролиферацию мезангиоцитов (см. ниже). На поверхности подоцитов есть рецепторы к белкам системы комплемента и антигенам, что свидетельствует об активном участии этих клеток в иммуновоспалительных реакциях.
Фильтрационный барьер
Все три названных компонента - эндотелий капилляров сосудистого клубочка, подоциты внутреннего листка капсулы и общую для них гломерулярную базальную мембрану - принято перечислять в составе фильтрационного барьера, через который из крови в полость капсулы фильтруются составные части плазмы крови, образующие первичную мочу. Если более внимательно проанализировать данную ситуацию, то к
данному перечислению необходимо внести некоторые уточнения; в этом случае состав собственно фильтрационного барьера будет выглядеть следующим образом:
фенестры и щели эндотелия капилляров;
3-слойная базальная мембрана;
щелевые диафрагмы подоцитов.
Примечание: избирательная проницаемость фильтрационного барьера может регулироваться некоторыми биологически активными веществами: например, повышению скорости фильтрации способствует предсердный натрийуретический фактор (пептид), а также ряд воздействий со стороны мезангиальных компонентов.
Мезангий
В сосудистых клубочках почечных телец в тех местах, куда между капиллярами не могут проникнуть цитоподии подоцитов (т.е. около 20% площади поверхности), находится мезангий - комплекс клеток (мезангиоцитов) и основного вещества (матрикса).
В большинстве руководств термин мезангий переводят как «межсосудистые клетки», хотя справедливости ради переведем правильно - брыжейка сосуда (в данном случае трофико-регуляторный компонент капиллярной петли сосудистого клубочка).
Выделяют три популяции мезангиоцитов: гладкомышечную, макрофагическую и транзиторную (моноциты из кровотока). Мезангиоциты гладкомышечного типа способны синтезировать все компоненты матрикса, а также сокращаться под влиянием ангиотензина, гистамина, вазопрессина и таким образом регулировать клубочковый кровоток, изменяя общую «геометрию» капиллярных петель. Мезангиоциты макрофагического типа несут на своей поверхности Fc-рецепторы и другие компоненты главного комплекса гистосовместимости 2-го типа, необходимые для фагоцитарной функции, а также la-антиген. Благодаря этому создается возможность для локальной реализации в клубочках иммуновоспалительной реакции (к сожалению, в некоторых случаях и аутоиммунной).
Основными компонентами матрикса являются адгезивный белок ламинин и коллаген, образующий тонкофибриллярную сеть. Вероятно, матрикс также участвует в фильтрации веществ из плазмы крови капилляров клубочка, хотя окончательно данный вопрос еще не решен.
Реабсорбция
Реабсорбция (обратный перенос веществ из первичной мочи в окружающий нефрон интерстиций и, в конечном итоге, в капилляры вторичной сосудистой сети) представляет собой весьма сложный каскад транспортных процессов, которые значительно различаются в разных отделах канальцевого аппарата нефрона. Различия в указанных процессах, естественно, обусловливают и различия в морфологии отделов нефрона. В функциональном отношении необходимо выделить по крайней мере 4 таких отдела: проксимальные извитые канальцы, петлю нефрона, дистальные извитые канальцы, и собирательные трубочки.
Проксимальные извитые канальцы
В проксимальных извитых канальцах происходит активная (т.е. за счѐт специально расходуемой энергии) реабсорбция значительной части воды и ионов, практически всей глюкозы и всех белков. Данная реабсорбция не регулируется гормонами и поэтому называется облигатной.
Белки переносятся путѐм пиноцитоза (из просвета канальца в цитоплазму канальцевых эпителиоцитов на их апикальных полюсах), который последовательно сменяется экзоцитозом (из цитоплазмы эпителиоцитов на базальных полюсах через базальную мембрану в капилляры вторичной сети). При этом многочисленные пиноцитозные пузырьки насыщают всю цитоплазму эпителиоцитов и продвигаются по ней с помощью ориентированных микротрубочек (здесь уместно вспомнить о тубулин-кинезиновом хемо-механическом преобразователе - см. лекции по цитологии). Существует мнение (наиболее распространенное), что поступающие в цитоплазму эпителиоцитов белки расщепляются в цитоплазме под влиянием лизосомальных протеолитических ферментов до аминокислот, которые затем транспортируются в кровь перитубулярных капилляров. По всей видимости, все же не все белки расщепляются до аминокислот - часть их (возможно, большая) переносится в неизмененном состоянии - ведь количество пиноцитозных структур на базальном полюсе клеток почти такое же, как и на апикальном.
Глюкоза (как, впрочем, и некоторые другие моносахариды) всасывается путѐм симпорта (т.е. сопряжѐнного переноса) с ионами Na, поступающими в эпителиальную клетку по градиенту их концентрации через особые каналы. Эти каналы функционируют только при наличии в первичной моче одновременно и Na+, и определенного моносахарида. Другими словами, при отсутствии одного из компонентов (либо Na+, либо глюкозы) данный канал не срабатывает. Напомним, что указанные каналы работают по принципу облегченной диффузии, т.е. в соответствии с градиентом концентрации (в данном случае Na+) и без затрат энергии. Такая работа требует постоянного поддержания низкой внутриклеточной концентрации ионов Na+, а это обеспечивается за счѐт деятельности Na+-насоса (уже энергозависимого) на базальной поверхности эпителиальных клеток.
Реабсорбируемая вода проходит непосредственно через цитоплазму эпителиальных клеток (а не через промежутки между ними) с помощью водных каналов (так называемых аквапоринов). Работа этих каналов также связана с градиентом напряжения воды в трех взаимосвязанных «отсеках» - просвете канальца, цитоплазме эпителиоцита и крови перитубулярной капиллярной сети.
В соответствии с функциональным профилем эпителиоцитов общая структура проксимальных извитых канальцев имеет характерный вид. В частности, канальцевый эпителий достигает максимальной высоты среди всех канальцев нефрона, что связано с высокой реабсорбционной активностью. Поверхность клеток покрыта щеточной каемкой, обусловленной протеканием начальных стадий пиноцитоза и необходимостью увеличения контактной поверхности мембран, насыщенных транспортными структурами (например, каналами). Цитоплазма клеток насыщена пиноцитозными пузырьками и лизосомами, что отражает активный перенос белков и придает клеткам «пенистый» вид, а всей цитоплазме - выраженную оксифилию. В своей базальной части клетки имеют исчерченность - базальный лабиринт, образованный внутренними складками цитолеммы (активный экзоцитоз белков) и расположенными между ними митохондриями (энергообеспечение Na+, К+, Са++ и других насосов). В прямой части проксимального канальца, кроме того, в его просвет секретируются некоторые органические продукты - креатинин и др.
Петля нефрона
Петля Генле состоит из тонкого канальца и прямого дистального канальца. В коротких и промежуточных нефронах тонкий каналец имеет только нисходящую часть, а в юкстамедуллярных нефронах - также длинную восходящую часть, которая переходит в прямой (толстый) дистальный каналец. Тонкий каналец имеет диаметр около 15 мкм. Стенка его образована плоскими эпителиоцитами. Такая морфология связана с функциональными особенностями данного отдела нефрона - здесь происходит пассивная реабсорбция воды. В нисходящих тонких канальцах цитоплазма эпителиоцитов светлая, бедная органеллами и ферментами. Реабсорбция воды реализуется на основе разности осмотического давления между мочой в канальцах и тканевой жидкостью интерстициальной ткани, в которой проходят сосуды мозгового вещества. Деятельность многочисленных водных каналов (аквапоринов) обеспечивает интенсивную реабсорбцию воды, которая, впрочем, не требует потребления энергии. Поэтому у клеток нет признаков высокой функциональной активности - щеточной каѐмки, оксифилии цитоплазмы, высокого содержания митохондрий, складчатости базальной плазмолеммы.
Дистальный извитой каналец
Здесь происходят два процесса, регулируемые гормонами и называемые поэтому факультативными:
активная реабсорбция оставшихся электролитов и
пассивная реабсорбция воды.
В частности, работает Na+,К+-канал по принципу - обмен 3 ионов Na+ (внутрь цитоплазмы эпителиоцита) на 2 иона К+ и 1 ион Н+ (из цитоплазмы в мочу). Деятельность канала, не требующего энергии, основана на градиенте концентрации Na+; поддержание постоянной низкой концентрации Na+ в цитоплазме обеспечивается работой Na+ ,К +-насосов, активность которых регулируется гормоном альдостероном. Важно отметить, что указанные насосы расположены не на базальном полюсе канальцевых эпителиоцитов (как в проксимальных канальцах), а на их боковых поверхностях. При этом из цитоплазмы Na+ откачивается в исключительно узкое интерстициальное пространство между эпителиальными клетками, благодаря чему даже при незначительном количестве молекул Na+ в нем удается достигнуть резкого повышения интерстициального осмотического давления. Под действием этого высокого давления вода реабсорбируется в интерстициальные щели между эпителиоцитами и затем вместе с натриевыми ионами увлекается в перитубулярные гемокапилляры. Данная реабсорбция регулируется антидиуретическим гормоном (АДГ), который понижает полимерность гиалуроновой кислоты в составе гликозаминогликанов интерстиция, тем самым увеличивая его гидрофильность и интенсифицируя глубину реабсорбции воды. Соблюдается простая схема: чем больше АДГ, тем меньше мочи и тем выше ее концентрированность.
Характерная морфология дистального канальца: он выстлан низким цилиндрическим эпителием, клетки которого лишены щеточной каемки, но имеют базальный лабиринт с высоким содержанием митохондрий (для энергообеспечения Na+K+-нacocoв).
Собирательные трубочки
Собирательные трубочки в верхней (корковой) части выстланы однослойным кубическим эпителием, а в нижней (мозговой) части — однослойным низким цилиндрическим эпителием. В эпителии различают светлые и темные клетки. Светлые клетки бедны органеллами, их цитоплазма образует внутренние складки. Темные клетки по своей ультраструктуре напоминают париетальные клетки желез желудка, секретирующие соляную кислоту. Кроме ионов хлора, данные клетки секретируют в мочу аммиак. В собирательных трубочках с помощью светлых клеток завершается пассивное обратное всасывание из мочи в кровь части воды. Кроме того, происходит подкисление мочи, что, вероятно, связано с секреторной деятельностью темных эпителиоцитов, выделяющих в просвет трубочек ионы водорода и аммония.
Реабсорбция воды в собирательных трубочках зависит от концентрации в крови антидиуретического гормона гипофиза. В его отсутствие стенка собирательных трубочек и конечных участков извитых дистальных канальцев непроницаема для воды, поэтому концентрация мочи не повышается, а ее количество не изменяется. В присутствии гормона стенки указанных канальцев становятся весьма проницаемыми для воды, которая выходит в гипертоническую среду интерстиция мозгового вещества (пассивно, путем осмоса по механизму, близкому к описанному в дистальных извитых канальцах) и затем в перитубулярные капилляры. В этом процессе важную роль играют прямые сосуды (сосудистые пучки), которые уносят воду, поступающую из собирательных трубочек. В результате по мере продвижения по собирательным трубочкам моча становится все более концентрированной и из организма выделяется гипертонической (вторичная моча).
Разнообразие морфологических типов эпителиоцитов в данном отделе почечных канальцев, а также увеличение объема цитоплазмы эпителиальных клеток отражает нарастание процессов третьей - секреторной - фазы мочеобразования.
Таким образом, расположенные в мозговом веществе канальцы нефронов (тонкие, прямые дистальные) и медуллярные отделы собирательных трубочек, участвующие в реабсорбции электролитов и воды в комплексе с гиперосмолярной интерстициальной тканью мозгового вещества и перитубулярными гемокапиллярами составляют противоточно-множительный аппарат почек. Именно этот аппарат обеспечивает концентрирование и уменьшение объема выделяемой мочи, что является механизмом для регуляции водно-солевого гомеостаза в организме.
Эндокринная система почек
Данная система участвует в регуляции кровообращения и мочеобразования в почках и оказывает влияние на общую гемодинамику и водно-солевой обмен в организме. Система включает 3 основных компонента: ренин-ангиотензин-альдостероновый, простагландиновый и калликреин-кининовый аппараты.
Ренин-ангиотензиновый аппарат
Он же - юкстагломерулярный аппарат (ЮГА), околоклубочковый. В ЮГА входят 3 компонента: плотное пятно, ЮГ клетки и ЮВ клетки Гурмагтига.
1. Плотное пятно (macula densa) - тот участок стенки дистального извитого канальца, который прилегает к почечному тельцу. Границы между клетками почти не видны, у клеток нет базальной исчерченности, но хорошо различается скопление гиперхромных ядер, расположенных на близком расстоянии (отчего это место и выглядит в виде плотного базофильного пятна). Клетки плотного пятна утрачивают способность к реабсорбции (не имея возможности взаимодействовать с пери¬тубулярными капиллярами), но подобно «натриевому рецептору» улавливают изменения содержания натрия в моче и воздействует на юкстагломерулярные клетки, секретирующие ренин. Таким образом, плотное пятно выполнет функции осморецептора.
2. Юкстагломерулярные клетки - находятся в стенке приносящей и выносящей артериол, образуя второй слой клеток, лежащий под эндотелием. Данные клетки по происхождению и локализации являются гладкими миоцитами, однако утрачивают функцию сокращения, перестраиваясь на секрецию гормона ренина. Являются крупными клетками овальной или полигональной формы, с крупными гранулами, содержащими ренин.
Секреция ренина стимулируется двумя факторами: 1) раздражением осморецептора (клеток плотного пятна) при нарастании концентрации Na+ и 2) раздражением барорецепторов в стенке приносящей и выносящей артериол (при снижении давления крови в их просвете).
3. Юкставаскулярные клетки (клетки Гурмагтига) - это клетки, расположенные в треугольном пространстве между двумя артериолами (приносящей и выносящей) и плотным пятном. Клетки имеют длинные отростки, контактирующие с другими клетками мезангия. По происхождению и локализации клетки Гурмагтига относятся к мезангиальным клеткам, формируя особую популяцию в их составе. В обычных условиях данные клетки вырабатывают фермент ангиотензиназу, который обусловливает инактивацию ангиотензина (см. ниже) и таким образом «противоборствует» деятельности ренин-ангиотензин-альдостеронового аппарата (также см. ниже). При некоторых обстоятельствах (например, при стрессе, повышенной физической нагрузке, а также при истощении длительно функционирующих юкстагломерулярных клеток) клетки Гурмагтига утрачивают свою антагонистичность; боле того, они сами «переключаются» на синтез ренина.
Ренин - представляет собой полипептид с ферментативной активностью. В крови он воздействует на неактивный пептид (вырабатываемый печенью) - ангиотензиноген, который в две стадии превращается в свою активную форму - ангиотензин II. Этот продукт, во-первых, повышает тонус миоцитов мелких сосудов и тем самым повышает давление, а во-вторых, стимулирует выделение альдостерона в коре надпочечников. Последнее же может усиливать выработку антидиуретического гормона. Таким образом, избыточная продукция ренина приводит не только к спазму мелких сосудов, но и к усилению реабсорбирующей функции самих почек. Происходящее увеличение объѐма плазмы крови в еще большей степени (наряду со спазмом сосудов) повышает давление крови.
Простагландиновый аппарат
По своему действию на почки простагландиновый аппарат является антагонистом ренин-ангиотензин-альдостеронового аппарата. Почки могут вырабатывать (из полиненасыщенных жирных кислот) гормоны простагландины - жирные кислоты, содержащие в своей структуре пятиуглеродный цикл. Группа этих веществ очень разнообразна - также как и вызываемые ими эффекты. Фракция «почечных» простагландинов оказывают сосудорасширяющее действие, увеличивают клубочковый кровоток, объем выделяемой мочи и экскрецию с ней ионов Na. Стимулами для выделения простагландинов в почках являются ишемия, повышение содержания ангиотензина, вазопрессина, кининов.
Синтез простагландинов в почках осуществляется двумя видами клеток мозгового вещества: светлыми клетками собирательных трубочек и интерстициальными клетками. Интерстициальные клетки находятся в строме мозговых пирамид. Своими отростками интерстициальные клетки оплетают с одной стороны - каналец петли Генле, а с другой стороны - кровеносный капилляр. В теле этих клеток находятся гранулы, содержащие простагландины.
Калликреин-кининовый аппарат
Данный аппарат обладает сильным сосудорасширяющим действием и повышает натрийурез и диурез путем угнетения реабсорбции Na и воды в канальцах нефронов. Кинины - это небольшие пептиды, которые образуются под влиянием ферментов калликреинов из белков-предшественников (кининогенов), содержащихся в плазме крови. В почках калликреины выявляются в клетках дистальных канальцев, и на их уровне происходит высвобождение кининов. Вероятно, свое действие кинины оказывают, стимулируя секрецию простагландинов.
Возрастные изменения
Возрастные особенности строения почек указывают на то, что выделительная система человека в постэмбриональном периоде продолжает свое развитие длительное время. Так, по толщине корковый слой у новорожденного составляет всего 4/5, а у взрослого - 1/3 мозгового вещества, однако при этом увеличение массы почечной ткани связано не с образованием новых, а с ростом и дифференцировкой уже существующих нефронов, которые в детском возрасте еще не полностью развиты. В почке ребенка обнаруживается большое число нефронов с мелкими нефункционирующими и слабодифференцированными клубочками. Толщина извитых канальцев нефронов у детей в среднем 18-36 мкм, тогда как у взрослого она достигает 40-60 мкм. Особенно резким изменениям с возрастом подвергается длина нефронов. Их рост продолжается вплоть до половой зрелости. Поэтому с возрастом, по мере того как увеличивается масса канальцев, количество клубочков на единицу поверхности почки уменьшается. Подсчитано, что у новорожденных на один и тот же объем почечной ткани приходится до 50 клубочков, у 8-10-месячных детей - 18-20 клубочков, а у взрослых - 4-6 клубочков.
МОЧЕВЫВОДЯЩИЕ ПУТИ
К мочевыводящим путям относятся почечные чашки (малые и большие), лоханки, мочеточники, мочевой пузырь и мочеиспускательный канал, который у мужчин одновременно выполняет функцию выведения из организма семенной жидкости и поэтому будет описан в лекции «Мужская половая система».
Строение стенок почечных чашек и лоханок, мочеточников и мочевого пузыря в общих чертах сходно. В них различают слизистую оболочку, состоящую из переходного эпителия и собственной пластинки, подслизистую основу, мышечную и наружную оболочки.
В стенке почечных чашек и почечных лоханок вслед за переходным эпителием располагается собственная пластинка слизистой оболочки, незаметно переходящая в соединительную ткань подслизистой основы. Мышечная оболочка состоит из тонких слоев спирально расположенных гладких миоцитов, однако вокруг сосочков почечных пирамид миоциты имеют циркулярное расположение. Наружная оболочка без резких границ переходит в соединительную ткань, окружающую крупные почечные сосуды.
Мочеточники обладают выраженной способностью к растяжению благодаря наличию в них глубоких продольных складок слизистой оболочки. В подслизистой основе нижней части мочеточников располагаются мелкие альвеолярно-трубчатые железы. Мышечная оболочка, образующая в верхней части мочеточников два, а в нижней части - три слоя, состоит из гладкомышечных пучков, охватывающих мочеточник в виде спиралей, идущих сверху вниз. Они являются продолжением мышечной оболочки почечных лоханок и внизу переходят в мышечную оболочку мочевого пузыря, имеющую также спиралевидное строение. Лишь в той части, где мочеточник проходит через стенку мочевого пузыря, пучки гладких мышечных клеток идут только в продольном направлении. Сокращаясь, они раскрывают отверстие мочеточника независимо от состояния гладких мышц мочевого пузыря.
Спиральная ориентация гладких миоцитов в мышечной оболочке соответствует представлению о порционном характере транспорта мочи из почечной лоханки по мочеточнику. Согласно этому представлению, мочеточник состоит из 3, реже из 2 или 4 секций - цистоидов, между которыми находятся сфинктеры. Роль сфинктеров выполняют расположенные в подслизистой и в мышечной оболочках кавернозноподобные образования из широких извивающихся сосудов. В зависимости от наполнения их кровью сфинктеры оказываются закрытыми или открытыми. Происходит это последовательно рефлекторным образом по мере наполнения секции мочой и повышения давления на рецепторы, находящиеся в стенке мочеточника. Благодаря этому моча поступает порциями из почечной лоханки в вышележащие, а из нее - в нижележащие секции мочеточника, затем в мочевой пузырь. Снаружи мочеточники покрыты соединительнотканной адвентициальной оболочкой.
Слизистая оболочка мочевого пузыря состоит из переходного эпителия и собственной пластинки. В ней мелкие кровеносные сосуды особенно близко подходят к эпителию. В спавшемся или умеренно растянутом состоянии слизистая оболочка мочевого пузыря имеет множество складок. Они отсутствуют в переднем отделе дна пузыря, где в него впадают мочеточники и выходит мочеиспускательный канал. Этот участок стенки мочевого пузыря, имеющий форму треугольника, лишен подслизистой основы, и его слизистая оболочка плотно сращена с мышечной оболочкой. Здесь в собственной пластинке слизистой оболочки заложены железы, подобные железам нижней части мочеточников.
Мышечная оболочка мочевого пузыря построена из трех нерезко отграниченных слоев, которые представляют собой систему спирально ориентированных и пересекающихся пучков гладкомышечных клеток. Гладкие миоциты часто напоминают по форме расщепленные на концах веретена. Прослойки соединительной ткани разделяют мышечную ткань в этой оболочке на отдельные крупные пучки. В шейке мочевого пузыря циркулярный слой формирует мышечный сфинктер.
Наружная оболочка на верхнезадней и боковых (отчасти) поверхностях мочевого пузыря образована типичной серозной оболочкой (висцеральной брюшиной); в остальных участках - типичной адвентициальной оболочкой.
Мужская половая система.
Мужские половые железы
Яички, или семенники (testes),—мужские гонады,в которых образуются мужскиеполовые клетки - сперматозоиды и мужской половой гормон — тестостерон.
Развитие
Развитие мужской и женской гонады начинается однотипно (т.н. индифферентная стадия) и тесно связано с развитием выделительной системы.
Различают три составные элемента развивающихся половых желез:
 гоноциты, дающие начало овогониям или сперматогониям; гоноциты, дающие начало овогониям или сперматогониям;
 производные мезодермального целомического эпителия – будущие эпителиальные элементы половых желез; производные мезодермального целомического эпителия – будущие эпителиальные элементы половых желез;
мезенхимная ткань – соединительнотканные и гладкомышечные элементы гонад.
На медиальной стороне первичной почки (мезонефроса) образуются гребневидные утолщения – половые валики или будущие гонады. Целомический эпителий дает начало половым шнурам,врастающим в половые валики.В дальнейшем половые шнурыразвиваются в семенные канальцы (tubuli seminiferi), но часть этих шнуров превращается в сеть семенника (rete testis).
В постнатальном периоде в семенных канальцах гоноциты размножаются, а эпителий половых шнуров сохраняется в качестве поддерживающих клеток. В тех же половых шнурах, из которых развивается сеть яичка, гоноциты постепенно редуцируются.
Канальцы сети яичка,приближаясь к белочной оболочке средостения,сливаются в выносящие канальцы (ductuli efferentes),формирующиеся путем перестройкиэпителиальной выстилки канальцев первичной почки. Выносящие канальцы яичка, собираясь, переходят далее в канал придатка яичка (ductus epididymis), проксимальный отдел которого, многократно извиваясь, образует придаток семенника (epididymis), тогда как его дистальная часть становится семявыносящим протоком (ductus defferes).
Отводящие пути мужской половой системы развиваются из мезонефральных протоков, или Вольфовых каналов.
Парамезонефральный (мюллеров) проток в мужском организме атрофируется и сохраняется только верхний конец и нижний, превращающийся в предстательную (мужскую) маточку (utriculus prostaticus), располагающуюся у взрослого мужчины в толще предстательной железы у места впадения семявыносящего протока в мочеиспускательный канал. Предстательная железа и семенные пузырьки развиваются как выросты мочеполового синуса. После22-й недели гоноциты превращаются всперматогонии, при этом они утрачивают гликоген и высокую активность щелочной фосфатазы.
В онтогенезе эндокринная функция яичка устанавливается раньше, чем генеративная. Мужской половой гормон — тестостерон начинает вырабатывать у зародыша человека ГПД - ген половой детерминации приблизительно с 8—10-й недели внутриутробного периода. В зародышевом яичке еще до начала биосинтеза тестостерона образуются поочередно несколько белковых (пептидных) андрогенов, принадлежащих к ингибинам. Первым на стадии индифферентной гонады появляется гормон, под влиянием которого происходит редукция парамезонефрального протока, и с этого момента индифферентный зачаток половой системы дифференцируется по мужскому типу.
К середине эмбриогенеза, когда в семенных канальцах развивающегося яичка уже дифференцируется эпителиосперматогенный слой, а в просвете в большом количестве скапливаются гоноциты, начинается выработка второго ингибина, который, с одной стороны, угнетает размножение гоноцитов и вызывает их разрушение, а с другой — одновременно ингибирует секрецию ФСГ аденогипофизом. Этот пептид вырабатывается эпителием сети яичка.
Наконец, по мере приближения к пренатальному периоду эмбриогенеза у плода мужского пола появляется ингибин, который, сохраняя способность избирательно угнетать секрецию ФСГ, действует на соответствующие центры гипоталамуса.
Строение
Снаружи большая часть семенника покрыта серозной оболочкой — брюшиной, под которой располагается плотная соединительнотканная белочная оболочка, (tunica albuginea). На заднем крае яичка она утолщается, формируя средостение (mediastinum testis), от которого в глубь железы отходят соединительнотканные перегородки (septula testis), разделяющие железу на дольки (около 250 долек). В каждой дольке находится от 1 до 4 извитых семенных канальцев (tubuli seminiferi convoluti). Каждый семенной каналец имеет диаметр от 150 до 250 мкм и длину от 30 до 70 см. Приближаясь к средостению, канальцы (300—450 в каждом семеннике), сливаются и становятся прямыми, а в толще средостения соединяются с канальцами сети семенника. Из этой сети выходят (около 10) выносящие (эфферентные) канальцы (ductuli efferens), впадающих в проток придатка (ductus epididymis).
Внутреннюю выстилку семенных канальцев образует эпителиосперматогенный слой (или так называемый сперматогенный эпителий), расположенный на базальной мембране.
Кнаружи от эпителиосперматогенного слоя семенного канальца расположена собственная оболочка (tunica propria), состоящая из трех слоев:базального слоя (stratum basale), миоидного слоя (stratum myoideum) и волокнистого слоя (stratum fibrosum). Базальный слой (внутренний волокнистый слой), расположенный между двумя базальными мембранами (сперматогенного эпителия и миоидных клеток), состоит из сети коллагеновых волокон. Миоидный слой образован миоидными клетками, содержащими актиновые филаменты. Миоидные клетки обеспечивают ритмические сокращения стенки канальцев. Наружный волокнистый слой состоит из двух частей. Непосредственно к миоидному слою примыкает неклеточный слой, образованный базальной мембраной миоидных клеток и коллагеновыми волокнами. За ними расположен слой, состоящий из фибробластоподобных клеток, прилежащий к базальной мембране эндотелиоцитов гемокапилляра.
В соединительной ткани между семенными канальцами расположены гемокапилляры и лимфокапилляры, обеспечивающие обмен веществ между кровью и сперматогенным эпителием. Избирательность поступления веществ из крови в сперматогенный эпителий и различия в химическом составе плазмы крови и жидкости из семенных канальцев позволили сформулировать представление о гематотестикулярном барьере. Гематотестикулярным барьером называется совокупность структур, располагающихся между просветами капилляров и семенных канальцев.
Эпителиосперматогенный слой (epithelium spermatogenicum)имеет две основныхпопуляции клеток:
сперматогенные клетки (cellulae spermatogenicae), находящиеся на различных стадиях дифференцировки (стволовые клетки, сперматогонии, сперматоциты, сперматиды и сперматозоиды) и
поддерживающие клетки, или сустентоциты (epitheliocytus sustentans), или же клетки Сертоли.
Обе популяции клеток находятся в тесной морфофункциональной связи. Поддерживающие клетки (клетки Сертоли) лежат на базальной мембране, имеют пирамидальную форму и достигают своей вершиной просвета извитого семенного канальца. Ядра их имеют неправильную форму с инвагинациями, трехчленное ядрышко (ядрышко и две группы околоядрышкового хроматина). В цитоплазме особенно хорошо развита агранулярная эндоплазматическая сеть, аппарат Гольджи. Встречаются также микротрубочки, микрофиламенты, лизосомы и особые кристаллоидные включения. Обнаруживаются включения липидов, углеводов, липофусцина. На боковых поверхностях сустентоцитов образуются бухтообразные углубления, в которых располагаются дифференцирующиеся сперматогонии, сперматоциты и сперматиды.
Между соседними поддерживающими клетками формируются зоны плотных контактов, которые подразделяют сперматогенный эпителий на два отдела — наружный базальный и внутренний адлюминальный.
В базальном отделе расположены сперматогонии, имеющие максимальный доступ к питательным веществам, поступающим из кровеносных капилляров.
В адлюминальном отделе находятся сперматоциты на стадии мейоза, а также сперматиды и сперматозоиды, которые не имеют доступа к тканевой жидкости и получают питательные вещества непосредственно от клеток Сертоли.
Поддерживающие эпителиоциты создают микросреду, необходимую для дифференцирующихся половых клеток, изолируют формирующиеся половые клетки от токсических веществ и различных антигенов, препятствуют развитию иммунных реакций. Таким образом сустентоциты – основной компонент гематотестикулярного барьера. Кроме того, сустентоциты способны к фагоцитозу дегенерирующих половых клеток и последующему лизису их с помощью своего лизосомального аппарата.
Клетки Сертоли синтезируют андрогенсвязывающий белок (АСБ), который транспортирует мужской половой гормон к сперматидам. Секреция АСБ усиливается под влиянием ФСГ аденогипофиза. Поддерживающие эпителиоциты имеют поверхностные рецепторы для ФСГ, а также рецепторы для тестостерона и его метаболитов.
Различают два вида поддерживающих клеток — светлые и темные. Светлые поддерживающие клетки продуцируют ингибин – фактор, тормозящий секрецию ФСГ аденогипофизом. Темные поддерживающие клетки продуцируют фактор, стимулирующий деление сперматогенных клеток.
Генеративная функция. Сперматогенез.
Образование мужских половых клеток (сперматогенез) протекает в извитых семенных канальцах и включает 4 последовательные стадии или фазы: размножение, рост, созревание и формирование.
Начальной фазой сперматогенеза является размножение сперматогоний, занимающих наиболее периферическое (базальное) положение в сперматогенном эпителии. Согласно современным представлениям, среди сперматогоний можно выделить два типа клеток: 1) стволовые сперматогонии типа А, которые подразделяются на две субпопуляции: долгоживущие, резервные стволовые клетки и быстро обновляющиеся полустволовые клетки, которые делятся один раз в течение цикла сперматогенного эпителия, 2) дифференцирующиеся сперматогоний типа А и типа В.
Стволовые клетки расположены в базальной части канальца изолированно от других сперматогоний. Морфологически в популяции стволовых А-сперматогоний различают светлые и темные клетки. Для обоих клеток характерно преобладание в ядрах деконденсированного хроматина и расположение ядрышек около ядерной оболочки. Однако в темных клетках типа А степень конденсации хроматина большая, чем в светлых клетках. Темные клетки относят к «резервным» медленно обновляющимся стволовым клеткам, а светлые — к полустволовым быстро обновляющимся клеткам. Для стволовых клеток характерно наличие овальных ядер с диффузно распределенным хроматином, одного или двух ядрышек, большое содержание в цитоплазме рибосом и полисом, малое количество других органелл.
|
|
|
 Скачать 159.82 Kb.
Скачать 159.82 Kb.