Лимбическая система мозга
 Скачать 0.52 Mb. Скачать 0.52 Mb.
|

|
Роль лимбической системы в интегративной деятельности мозга. Современные представления о локализации функций в коре больших полушарий. Методы исследования ЦНС. Лимбическая система мозга 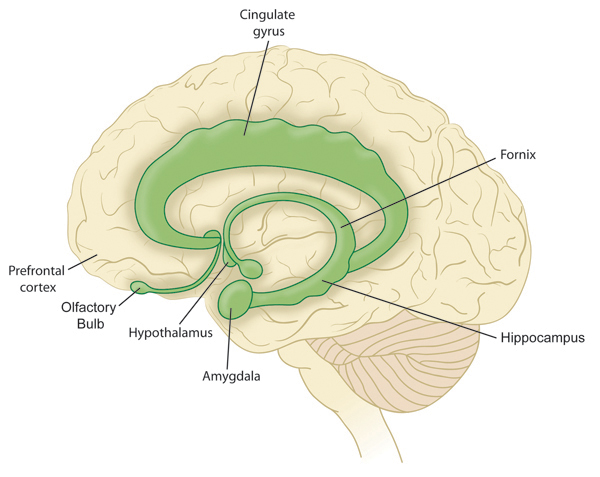 Целостная поведенческая реакция осуществляется благодаря объединению деятельности многих структур головного мозга. Лимбическая система мозга — это совокупность функционально, морфологически и филогенетически связанных между собой различных образований мозга, выполняющих следующие функции: - мотивационно-эмоциональную; - памяти и обучения; - регуляции цикла «сон-бодрствование», - репродуктивного и сексуального поведения. Корковые структуры лимбической системы: · древняя кора- препериформная, периамигдалярная, диагональная кора; грушевидная доля (находится в базальной части височной доли), обонятельная луковица, обонятельный бугорок, прозрачная перегородка · старая кора- гиппокамп, поясная извилина (лимбическая кора), область, лежащая возле поясной извилины (субикулум), орбитальная часть орбитофронтальной коры; · новая кора - лобная и теменная доли коры. Подкорковые стуктуры: гипоталамус, миндалина, ядра перегородки, передние ядра таламуса, сосцевидные тела таламуса, ретикулярная формация среднего мозга, центральное серое вещество среднего мозга. Связи лимбической системы: Все многочисленные формирования лимбической коры кольцеобразно охватывают основание переднего мозга и являются своеобразной границей между новой корой и стволовой частью мозга. Лимбическая система характеризуется обилием двусторонних связей с другими отделами мозга и внутри самой системы. Так, например, установлено наличие мощных связей лимбической системы с гипоталамусом. Через гипоталамус и сосцевидные тела лимбическая система соединена с центральным серым веществом и ретикулярной формацией среднего мозга. Прямых связей между лимбической системой и остальными частями коры очень мало (трудно сдерживать свои эмоции). Афферентные: импульсы от коры (чувствительных, ассоциативных, двигательных зон) поступают в лимбическую систему через передние ядра таламуса (исключение связь с лобной корой), от обонятельных луковиц, от гипоталамуса, от вегетативных ядер ствола. Эфферентные пути: к вегетативным центрам гипоталамуса, от них к гипофизу, к базальным ганглиям, вегетативной нервной системе, ретикулярной формации. При раздражении структур лимбической системы наблюдается изменение деятельности внутренних органов и сосудов, но эти реакции носят фазный характер (легко возникают и быстро проходят). Образование лимбической системы оказывают модулирующие влияние, приспосабливая деятельность внутренних органов для осуществления поведенческих реакций. Морфофункциональная организация. Особенностью лимбической системы является то, что между ее структурами имеются простые двусторонние связи и сложные пути, образующие множество замкнутых кругов. Такая организация создает условия для длительного циркулирования одного и того же возбуждения в системе и тем самым для сохранения в ней единого состояния и навязывание этого состояния другим системам мозга. В настоящее время хорошо известны связи между структурами мозга, организующие круги, имеющие свою функциональную специфику. Большой круг Папеца: поясная извилина моносинаптически связана с гиппокампом, он через свод с мамиллярными телами гипоталамуса, который через передние ядра таламуса с поясной извилиной. 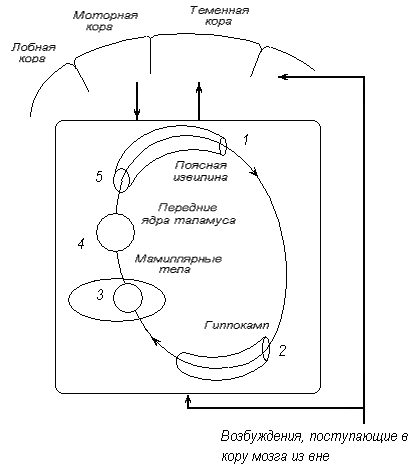 Этот круг имеет отношение к памяти и процессам обучения. Другой круг, круг Наута (миндалевидное тело - гипоталамус - мезенцефальные структуры - миндалевидное тело) регулирует агрессивно-оборонительные, пищевые и сексуальные формы поведения. Оба эти круга связаны через гипоталамус 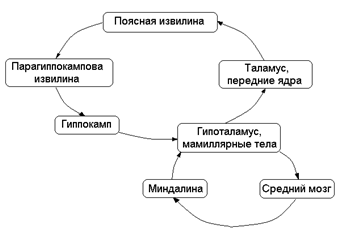 Считается, что образная память формируется кортико-лимбико-таламо-кортикальным кругом. Круги разного функционального назначения связывают лимбическую систему со многими структурами центральной нервной системы, что позволяет последней реализовать функции, специфика которых определяется включенной дополнительной структурой. Например, включение хвостатого ядра в один из кругов лимбической системы определяет ее участие в организации тормозных процессов высшей нервной деятельности. Большое количество связей в лимбической системе, своеобразное круговое взаимодействие ее структур создают благоприятные условия для реверберации возбуждения по коротким и длинным кругам. Это, с одной стороны, обеспечивает функциональное взаимодействие частей лимбической системы, а с другой – создает условия для запоминания. Обилие связей лимбических структур с образованиями центральной нервной системы затрудняет выделение функций мозга, в которых она не принимала бы участия. Так, лимбическая система имеет отношение к регулированию уровня реакции вегетативного и соматического отделов центральной нервной системы при эмоционально-мотивационной деятельности, регулированию уровня внимания, восприятия, воспроизведения эмоционально значимой информации. Она определяет выбор и реализацию адаптивных форм поведения, динамику врожденных форм поведения, поддержание гомеостаза, генеративных процессов. Наконец, она обеспечивает создание эмоционального фона, формирование и реализацию процессов высшей нервной деятельности. Нужно отметить, что древняя и старая кора лимбической системы имеет прямое отношение к обонятельной функции. В свою очередь обонятельная сенсорная система, как самая древняя из них, является неспецифическим активатором всех видов деятельности коры головного мозга. Часто лимбическую систему называют висцеральным мозгом, то есть структурой центральной нервной системы, участвующей в регуляции деятельности внутренних органов. И действительно, миндалевидные тела, перегородка, обонятельный мозг при их возбуждении изменяют активность вегетативных систем организма в соответствии с условиями окружающей среды. Роль гипоталамуса в лимбической системе Важнейшая функция лимбической системы – управление работой внутренних органов, желез и сосудов. Здесь главенствует гипоталамус. По сравнению с обширными полушариями он кажется совсем крохотным. Однако нарушение работы центров, сосредоточенных на этом «пятачке», влечет за собой всевозможные недуги: ожирение и бессонницу, диабет и лихорадку, гипертонию и половое бессилие. Еще из гипоталамуса мозг «дирижирует» всей эндокринной системой. Не удивительно, что эмоциональные бури, прокатываясь по лимбической системе, так сильно сказываются на состоянии нашего организма. Одной из важных и характерных функций гопоталамуса является широкое представительство «химической кухни», которая богата регуляторами и поэтому чувствительна к воздействию психоактивных веществ (типа эндорфинов). Поэтому в гипоталамусе имеются зоны, раздражение которых вызвает желание повторить это воздействие (реакция самораздражения), либо наоборот – никогда не повторять это воздействие. Другими словами, в этой области мозга есть зоны при активации которых возникает положительно окрашенное состояние или реакция обратного знака (негативное состояние). В определенном возрасте – в раннем детстве и в подростковом периоде – в лимбической системе происходит импринтинг, или запечатление образов для слепого подражания, для запуска автоматических программ поведения. В эти периоды из-за обостренной эмоциональности в памяти больше откладывается не логическая суть событий, а связанные с ними переживания. И ключом, позволяющим вновь включить эти чувства и побуждения – совершенно бессознательно, обычно становится химический сигнал: вкус, запах или дурманящее вещество. Так, иногда если пахнет хлоркой или краской, какая-то тоска вдруг сожмет сердце и что-то припомнится, в ушах отзовется детский визг, а язык будто обожжет вкус кефира или творога – того, что ел в детстве. Гиппокамп. Корковая структура – гиппокамп (hippocampus) расположен в глубине височных долей мозга и является основной структурой лимбической системы. Морфологически гиппокамп представлен стереотипно повторяющимися модулями, связанными между собой и с другими структурами. Строение. Особенности цитоархитектоники гиппокампа и его пирамидного слоя послужили основанием для его деления на 4 основных поля, которые ориентированы в виде полос в медиолатеральном направлении. Эти полосы не прерываясь тянутся от дорсального, переходя на задний и заканчиваясь в вентральном гиппокампе. 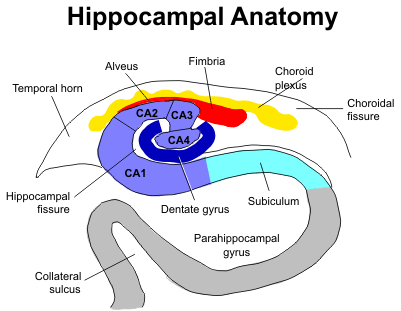 Модульное строение гиппокампа обусловливает его способность генерировать высокоамплитудную ритмическую активность. Связь модулей в гиппокампе создает условие для циркуляции активности при обучении. При этом возрастает амплитуда синаптических потенциалов, увеличиваются нейросекреция клеток гиппокампа, возрастает число шипиков на дендритах его нейронов, что свидетельствует о переходе потенциальных синапсов в активные. Многочисленные связи гиппокампа со структурами как лимбической системы, так и других отделов мозга определяют его полифункциональность. Основой функционирования нейронных ансамблей гиппокампа считают глутаматергическую нейромедиаторную систему, поскольку и пирамидные нейроны, и гранулярные клетки зубчатой фасции являются глутаматергическими. Разные сенсорные проекционные зоны в гиппокампе перекрываются. Это обусловлено тем, что большинство нейронов гиппокампа характеризуются полисенсорностью, то есть способностью реагировать на световые, звуковые и другие виды раздражений. Нейроны гиппокампа отличаются выраженной фоновой активностью. В ответ на сенсорное раздражение реагирует до 60% нейронов гиппокампа. Особенность строения гиппокампа, его взаимосвязанные модули обусловливают цикл генерирования возбуждения в нем, что выражается в длительной реакции нейронов (до 10-12 с) в ответ на однократный короткий стимул. Гиппокамп и память. Так какую же функциональную нагрузку несет на себе гиппокамп – этот важнейший отдел лимбического мозга? Оказалось, что этот отдел мозга принимает непосредственное участие в процессах кратковременной памяти. Благодаря гиппокампу мы запоминаем то, что «важно», а перед этим – замечаем все «новое». Информация, получившая эмоциональную окраску, запоминается не в пример лучше «мертвых фактов», которые, образно выражаясь «в один гиппокамп влетают – в другой вылетают...». А при сильных негативных переживаниях наблюдается обратный процесс: стирание следов памяти, то есть – амнезия. Повреждение гиппокампа ведет к снижению инициативности, замедлению скорости основных нервных процессов. Однако главным симптомом выпадения функций гиппокампа является утеря способности к обучению. Так, например, у нескольких человек было проведено двустороннее хирургическое удаление частей гиппокампа с целью лечения эпилепсии. Эти люди могли удовлетворительно вспоминать практически все ранее полученные знания. Однако при этом они не могли приобретать никаких новых сведений, основанных на вербальных сигналах. Действительно, обычно им не удается запомнить даже имена людей, с которыми они контактируют каждый день. Тем не менее, люди без гиппокампа на короткий период времени могут запомнить то, что происходит в процессе их текущей деятельности. Следовательно, они способны к кратковременной памяти, сохраняющейся в течение от нескольких секунд до 1-2 мин, хотя возможность запоминать что-либо на больший срок у них практически отсутствует, то есть имеет место антероградная амнезия. Гиппокамп, по-видимому, очень рано в эволюционном развитии мозга стал нервным механизмом для принятия критических решений, определяя значение входящих сенсорных сигналов. После установления этой способности принимать критические решения, остальная часть мозга, вероятно, также начинает обращаться к гиппокампу. Следовательно, если гиппокамп «говорит» субъекту, что данный нервный сигнал важен, он непременно должен быть зафиксирован в памяти. Гиппокамп – это отдел мозга, благодаря которому человек быстро перестает реагировать на индифферентные стимулы, как это бывает при ориентировочном рефлексе, но любое сенсорное переживание, вызывающее чувство удовольствия или боль – успешно запоминается. Возникает вопрос, какой механизм лежит в основе этого процесса? Предполагают, что гиппокамп обеспечивает возбуждение, переводящее кратковременную память в долговременную. Видимо, он передает некий сигнал или сигналы, которые заставляют разум повторять новую информацию до тех пор, пока не осуществится ее постоянное хранение. Каков бы ни был механизм, без гиппокампа консолидация долговременной памяти вербального или символического типа является слабой или не происходит совсем. Миндалевидный комплекс. Амигдала –это сложный комплекс ядер головного мозга, относящийся к базальным ядрам (архистриатум) и участвующий в осуществлении корригирующего влияния на деятельность стволовых образований мозга. Функции миндалевидного комплекса связаны с обеспечением оборонительного поведения, вегетативными, двигательными, эмоциональными реакциями, мотивацией условнорефлекторного поведения. Электрическая активность нейронов миндалин характеризуется разночастотными колебаниями. Фоновые ритмы могут коррелировать с ритмом дыхания и сердечных сокращений. Реакция ядра на внешние раздражения длится, как правило, до 85 мс, то есть значительно меньше, чем реакция на подобные же раздражения новой коры. Структуры ядер миндалины реагируют на зрительные, слуховые, интероцептивные, обонятельные, кожные раздражения, причем все эти раздражения вызывают изменение активности любого из ядер миндалины, то есть ядра миндалины полисенсорны. Нейроны имеют хорошо выраженную спонтанную активность, которая может быть усилена или заторможена сенсорными раздражениями. Многие нейроны полимодальны и полисенсорны. Раздражение ядер миндалевидного тела создает выраженный парасимпатический эффект в деятельности сердечнососудистой, дыхательной систем, приводит к понижению (редко к повышению) кровяного давления, урежению частоты сердечных сокращений, нарушению проведения возбуждения в проводящей системе сердца, возникновению аритмий и экстрасистолий. Сосудистый тонус может не изменяться. Урежение ритма сердечных сокращений при воздействии на миндалины отличается длительным скрытым периодом и имеет продолжительное последействие. Раздражение ядер миндалины вызывает угнетение дыхания, иногда кашлевую реакцию. Разнообразные эффекты раздражения миндалин обусловлены их связью с гипоталамусом, который регулирует работу внутренних органов. При искусственной активации миндалины появляются реакции принюхивания, облизывания, жевания, глотания, саливации, изменения перистальтики тонкой кишки, причемэффекты наступают с большим латентным периодом (до 30-45 с после раздражения). Животные с двусторонним поражением амигдалы отличались большим аппетитом, увеличением поглощаемой пищи и, как следствие – повышением веса. Повреждение миндалины у животных снижает адекватную подготовку вегетативной нервной системы к организации и реализации поведенческих реакций, приводит к гиперсексуальности, исчезновению страха, успокоению, неспособности к ярости и агрессии. Животные становятся доверчивыми. Например, обезьяны с поврежденной миндалиной спокойно подходят к гадюке, вызывавшей ранее у них ужас и бегство. Видимо, в случае повреждения миндалины исчезают некоторые врожденные безусловные рефлексы, реализующие память об опасности. При глубоких нарушениях в эмоционально-аффективной сфере при височной эпилепсии и психических заболеваниях, протекающих с психомоторными пароксизмами, феноменами беспокойства, страха, злости, с агрессивностью, сексуальными нарушениями, часто сочетающихся с различными висцеровегетативными проявлениями прибегают к оперативному лечению в виде разрушения ядер миндалевидного комплекса либо перерезкой внутриядерных путей или связей этой области с иными отделами мозга. Уточнение локализации ядер миндалевидного тела осуществляется электростимуляцией. Деструкции подвергают, как правило, либо вентролатеральную, либо дорсомедиальную часть миндалевидного комплекса. Операцию обычно осуществляют с двух сторон с интервалами между вмешательствами в 3-6 месяцев. Кора больших полушарий Высшим отделом центральной нервной системы является кора большого мозга (кора больших полушарий). Она обеспечивает совершенную организацию поведения животных, а равно и человека, на основе врожденных и приобретенных в онтогенезе функций. Морфофункциональная организация коры больших полушарий. Серое вещество, покрывающее борозды и извилины полушарий, называют корой или плащем большого мозга. Площадь поверхности коры в одном полушарии в среднем составляет для взрослого человека 22 м2. Причем на сводчатые (выпуклые) части извилин приходится 1/3, а на боковые и нижние стенки 2/3 всей площади коры. Одно полушарие содержит в среднем 10-14 млрд. нейронов (что составляет 70 % нейронов мозга) и 100-120 млрд. нейроглиальных клеток. В коре нейроны располагаются послойно: по верхнелатеральной поверхности в 6-7 слоев, медиальной – в 4-5, базальной – в 2-3 слоя. Поэтому толщина коры колеблется в пределах 1.5-5 мм. Глубокие борозды делят каждое полушарие большого мозга на лобную, височную, теменную, затылочную доли и островок. Островок расположен в глубине сильвиевой борозды и закрыт сверху частями лобной и теменной долей мозга. Как уже отмечалось, кора больших полушарий мозга делится на древнюю (paleocortex), старую (archicortex), новую (neocortex) и межуточную (meso-сortex) кору. Древняя кора составляет 0.6 % поверхности всей коры большого мозга. Это примитивно построенная корковая пластинка, нечетко отделяющаяся от подлежащих субкортикальных клеточных скоплений. Нейроны палеокортекса небольшие и на слои не дифференцированы. Они наряду с другими функциями, имеют отношение к обонянию и обеспечению взаимодействия систем мозга. Эта область представлена передним продырявленным пространством, периферической частью обонятельного мозга, пириформным полем, главная ее часть представлена периамигдалярной областью и миндалевидным комплексом, который тесно связан с прилежащей древней корой и его можно рассматривать как палеокортикальный компонент мозга. Характерной особенностью строения коры является ориентированное, горизонтально-вертикальное распределение ее нервных клеток по слоям и колонкам. Корковая структура отличается пространственно упорядоченным расположением функционирующих единиц и связей между ними. Прежде чем определять взаимное расположение и организацию слоев в коре головного мозга необходимо рассмотреть ее нейронное строение. Нейронный состав и распределение нейронов по слоям в разных областях коры различны, что позволило выделить в мозге человека более 50-ти цитоархитектонических полей (по Броудману). Разделение коры большого мозга на цитоархитектонические поля более четко формируется по мере совершенствования ее функции в филогенезе. Кора каждого из полушария головного мозга делится на несколько областей: затылочную, верхнюю и нижнюю теменные, постцентральную, область центральных извилин, прецентральную, лобную, височную, лимбическую, островковую. Каждая из них в соответствии с особенностями подразделяется на ряд полей, причем каждое поле имеет свое условное порядковое обозначение. Особенности строения и функций коры головного мозга. Кора больших полушарий имеет следующие морфофункциональные особенности: - цитоархитектоническое распределение коры больших полушарий с выделением по этому признаку полей; - многослойность расположения нейронов; - модульный принцип организации; - соматотопическое представительство периферических рецепторов; - экранность, то есть распределение периферической рецепции на плоскости нейронального поля коркового отдела сенсорной системы; - зависимость уровня корковой активности от влияния подкорковых структур и ретикулярной формации; - наличие представительства всех функций нижележащих структур центральной нервной системы; - перекрытие в коре большого мозга зон соседних периферических рецептивных полей; - наличие в специфических проекционных сенсорных и моторной систем, а также – вторичных и третичных полей с ассоциативными функциями; - наличие специализированных ассоциативных областей; - динамическую локализацию функций, выражающуюся в возможности компенсаций деятельности утраченных структур; - реципрокную функциональную взаимосвязь возбудительных и тормозных состояний; - способность к иррадиации возбуждения и торможения; - возможность длительного сохранения следов раздражения; - наличие специфической электрической активности. Цито- и миелоархитектоника коры головного мозга. Основной тип строения новой коры (рис. 3.76 А), с которым сравниваются все его участки, – это кора, состоящая из шести слоев, так называемая гомотипическая кора, по А.Бродману. Так, в двигательной зоне коры больших полушарий различают шесть основных слоев: молекулярный, наружный зернистый, пирамидный, внутренний зернистый, ганглионарный, слой полиморфных клеток. В первом (I) наружном молекулярном слое почти нет тел нейронов. Обнаруживаются единичные горизонтально ориентированные нейроны, тангенциальные ветвления нервных волокон нижележащих нейронов и глиальные клетки. Сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры больших полушарий. Второй (II) наружный зернистый слой содержит мелкие звездчатые и пирамидные нейроны размером около 10 мкм. Аксоны этих нейронов оканчиваются в III, IV и VI слоях коры головного мозга, а дендриты поднимаются в молекулярный слой. Третий (III) слой – это слой средних и крупных пирамидных нейронов, он самый широкий. Поскольку размеры клеток в этом слое неодинаковы, поэтому этот слой в большинстве корковых полей делят на подслои. Аксоны клеток III слоя образуют ассоциативные нервные волокна, идущие через белое вещество и соединяющие соседние участки коры, что видно на его миелоархитектонике (рис.3.76, А, справа). Четвертый (IV) внутренний зернистый слой содержит в основном мелкие звездчатые нейроны. В некоторых полях он разделяется на подслои, местами же резко истончается и даже вовсе исчезает. Этот слой сильно развит в зрительной и слуховой зонах коры головного мозга. Он состоит из сенсорных звездчатых нейронов, имеющих многочисленные ассоциативные связи с нейронами других типов. Аксоны этих клеток, разветвляются в пределах соседних как выше, так и ниже лежащих слоев коры. Здесь заканчиваются специфические таламокортикальные пути, то есть пути, начинающиеся от периферических рецепторов. Пятый (V) ганглионарный слой образован большими и гигантскими пирамидными нейронами (клетками Беца). В некоторых зонах мозга слой делится на подслои, в двигательной зоне он состоит из трех подслоев, средний из которых содержит гигантские пирамидные клетки Беца, достигающие в диаметре 120 мкм. Апикальные дендриты пирамидных нейронов достигают молекулярного слоя. Аксоны этих клеток идут в белое вещество, образуя комиссуральные и проекционные нервные волокна, и, прежде всего, пирамидные пути. Шестой (VI) слой полиморфных нейронов содержит также много эфферентных пирамидных нейронов. Кроме того, здесь есть веретенообразные нейроны. Дендриты нейронов VI слоя пронизывают всю толщину коры, достигая молекулярного слоя. Шестой слой нередко разделяют на два: поверхностный слой (VIa) и глубокий слой (VIb). Иногда глубокий слой (VIb) именуют седьмым (VII) слоем. Этот слой непосредственно переходит в белое вещество больших полушарий. Таким образом, в неокортекс содержит три типа нейронов, тела которых расположены в разных слоях. Это пирамидные клетки, клетки-зерна, или звездчатые клетки, и веретеновидные клетки. Первые два типа нейронов подразделяются еще на несколько видов. Функциональное значение: главные эфферентные слои - 5 и 6 слой, выходят за пределы КБП к нижележащим структурам, а 3 слой тоже эфферентный, но аксоны короткие и они осуществляют, в основном, внутрикортикальных связи. Главным афферентным путём к коре являются таламокортикальные проекции. Таламус играет важную роль, он пропускает в кору только биологически важные сигналы: - специфический таламокортикальный путь - идёт от специфических ядер таламуса к проекционным зонам коры; - неспецифический таламокортикальный путь - от неспецифических ядер таламуса, охватывает возбуждением всю кору больших полушарий. Это более древний путь, имеет важное значение в механизмах рассеянного внимания, бодрствования, выработки условных рефлексов.  Модульная организация коры головного мозга. Впервые на структурную упорядоченность нейронов коры головного мозга указал в 1957 году американский физиолог Вернон Маунткастл. Он привел физиологические доводы в пользу существования в соматосенсорной коре «колонок», составленных из функционально сходных нейронов. Такие колонки служили базой топической организации коры (рис.3.76 Б,В). Доказательством вертикальной колончатой организации коры головного мозга было следующее наблюдение: если микроэлектрод погружать перпендикулярно в кору, то на своем пути он встречает нейроны, реагирующие на один вид раздражения, если же микроэлектрод вводить горизонтально по коре, то он встречает нейроны, реагирующие на разные виды стимулов. В 1962 году Д.Хьюбел и Т.Визел выявили в зрительной коре мозга аналогичные колонки, которые они назвали ориентационными, поскольку все нейроны колонки, приходящую через толщу коры мозга «предпочитали» одинаковую ориентацию светлой полосы (рецепторного поля), на которую реагировали нейроны и впервые обозначили их для зрительной коры, как структурно-функциональные модули. Диаметр колонки составляет около 500 мкм и определяется она зоной распределения коллатералей восходящего афферентного таламокортикального волокна. Соседние колонки имеют взаимосвязи, организующие участки множества колонок в обеспечении той или иной реакции. Возбуждение одной из колонок приводит к торможению соседних. 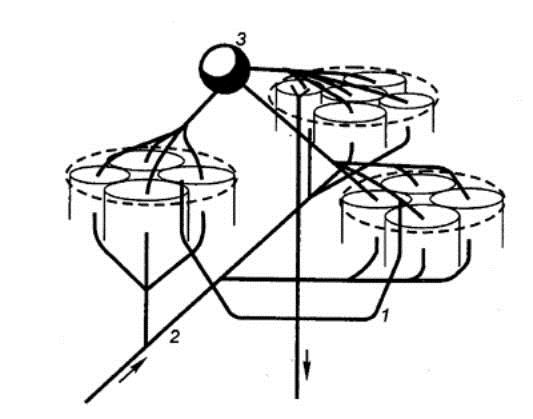 Рисунок. Функциональные объединения нейронных модулей. Одиночный структурный модуль имеет вид цилиндра; 4-6 модулей могут сгруппироваться в функциональные концентрические объединения, между которыми возможны как облегчающие, так и взаимотормозные отношения. Обширные функциональные объединения из нескольких концентрических модулей строятся на основе ветвлений аксонной коллатерали пирамидных нейронов (1), ветвлений таламического афферента (2) и ветвлений аксона корзинчатой клетки (3); стрелками показано направление восходящей и нисходящей импульсации. Топическая организация проекций. 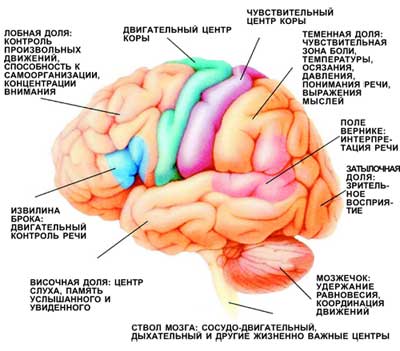 Сенсорная КБП - это центральные отделы всех сенсорных систем. Каждая сенсорная система имеет несколько сенсорных зон. -первичная зона - это центральный отдел ядер анализаторов, в них оканчивается сенсорные проекционные пути, имеет четкую соматотопическую организацию, благодаря им происходит первичный корковый анализ определённой сенсорной информации, при поражении этих зон может быть расстройства чувствительной и двигательной функции, проявляющиеся, например, в психической слепоте (человек видит предметы, а проанализировать свои ощущения не может); -вторичная зона - периферический отдел ядер анализаторов, расположено вокруг первичной зоны, характеризуется тем, что в ней идет дальнейший анализ сенсорной информации, отличие в том, что нейроны отвечают на раздражение различных модальностей, цвет и звук, свет и тактильные раздражение, благодаря им осуществляется взаимоотношение между анализаторами различных модальностей - процесс восприятия внешней среды - синтетическая деятельность мозга; -третичная зона (ассоциативная) - зона взаимного перекрытия анализаторов, особенно развита у человека, в ней развиты переключающие нейроны 2 и 3 слоев, благодаря которым осуществляется выработка условных рефлексов, участвует в организации интеллектуальной деятельности мозга, нейроны обладают высокой конвергентной емкостью, на одном может сходиться информация со многих зон, при поражении нарушается аналитико-синтетическая деятельность, речь, исчезают практические навыки, навык письма, целенаправленное (адаптивное) поведение. Ассоциативные области мозга у человека наиболее выражены в лобной, теменной и височной долях. Каждая проекционная область коры окружена ассоциативными областями. Нейроны этих областей чаще полисенсорны, обладают большими способностями к обучению. Так, в ассоциативном зрительном поле 18 число нейронов, «обучающихся» условнорефлекторной реакции на сигнал, составляет более 60% от числа фоновоактивных нейронов. Для сравнения: таких нейронов в проекционном поле 17 всего 10-12%. Полисенсорность нейронов ассоциативной области коры обеспечивает их участие в интеграции сенсорной информации, взаимодействие сенсорных и моторных областей коры. В теменной ассоциативной области коры формируются субъективные представления об окружающем пространстве, о нашем теле. Это становится возможным благодаря сопоставлению соматосенсорной, проприоцептивной и зрительной информации. Первой и наиболее характерной чертой ассоциативных областей коры является полисенсорность их нейронов, причем сюда поступает не первичная, а достаточно обработанная информация с выделением биологической значимости сигнала. Это позволяет формировать программу целенаправленного поведенческого акта. Вторая особенность ассоциативной области коры заключается в способности к пластическим перестройкам в зависимости от значимости поступающей сенсорной информации. Третья особенность ассоциативной области коры проявляется в длительном хранении следов сенсорных воздействий. Разрушение ассоциативной области коры приводит к грубым нарушениям обучения, памяти. Моторная КБП В моторной коре различают первичную моторную кору, а также премоторную и дополнительную моторную области 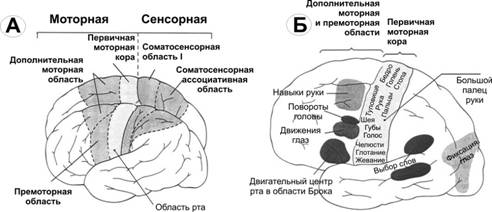 Первичная моторная кора. На рисунке дана грубая топография представительства различных мышц начиная от мышц лица (вблизи от сильвиевой борозды) и кончая мышцами ноги (область в глубине продольной щели). В первичной моторной коре картированы участки, стимуляция которых вызывает сокращения отдельных мышц, но чаще возбуждаются мышечные группы. · Премоторная область располагается кпереди от первичной моторной коры, её топографическая организация подобна организации первичной коры: наиболее латерально располагаются зоны лица и рта, далее вверх к продольной щели — кисть, рука, туловище и нижние конечности. В премоторной области генерируются сложные спектры движений (например, движения плеча, руки, особенно кисти). Кисть точно ориентирована для выполнения специфической работы. Чтобы достигнуть такого результата, передняя часть премоторной области формирует «двигательный образ» всех мышечных движений, которые должны осуществляться. Затем этот образ возбуждает каждый последовательный спектр мышечной активности в задней премоторной коре, требующийся для достижения «образа». Задняя премоторная кора посылает сигналы либо непосредственно в первичную моторную кору, возбуждая в итоге различные группы мышц, либо (чаще) через базальные ядра к первичной коре через таламус. Таким образом, премоторная кора, базальные ядра, таламус образуют комплексную систему контроля и координации спектра мышечной активности тела. · Дополнительная моторная область располагается в продольной щели и функционирует совместно с премоторной областью, обеспечивая движения, поддерживающие осанку, фиксацию движений различных сегментов тела, позиционные движения головы и глаз и базу для тонкого моторного контроля кистей рук премоторной областью и первичной моторной корой. · Специализированные области. На рисунке показано несколько относящихся к двигательной функции специализированных центров (например, центры формирования речи, произвольного движения глаз, вращения головы, целевого движения кисти) коры больших полушарий, впервые обнаруженных нейрохирургами (Пенфилд и Джаспер). Центр формирования речи. Непосредственно перед первичной моторной корой выше сильвиевой борозды располагается участок коры, называемый областью Брока. Повреждение этой области не лишает человека способности произносить звуки, но он теряет способность к осмысленному произнесению слов. В лучшем случае человек способен произносить слова «да» или «нет». Рядом расположенные корковые области управляют дыхательной активацией голосовых связок и движениями рта и языка во время разговора. Таким образом, формирование речи нейронами премоторной области является комплексным процессом, вовлекающим в активность прилежащие области коры. Центр произвольного движения глаз. Над областью Брока располагается участок, контролирующий произвольные движения глаз. Повреждение этого участка лишает человека способности смещать глаза в направлении различных объектов. Центр вращения головы находится рядом с центром, контролирующим движения глаз, этот центр направляет голову в сторону объектов. Центр целевого движения кисти лежит кпереди от первичной моторной коры. Повреждение этого центра делает движения кисти некоординированными и бессмысленными (моторная апраксия). Все речевые анализаторы закладываться в обоих полушариях, но развиваться только с одной стороны (у правшей – слева, у левшей – справа. Речедвигательный центр Брока — расположен в нижней части лобных извилин (поле 44) — это двигательный центр мышц языка. При поражении моторного центра речи развивается моторная афазия — в этом случае человек понимает речь, но сам, увы, говорить не может. Сенсорный центр Вернике — расположен в височной зоне в задних отделах верхней височной извилины (поля 22, 37, 42 левого полушария) — связан с восприятием устной речи. Задача этого центра – распознавание и хранение устной речи, как собственной, так и чужой. При поражении возникает сенсорная афазия — человек не воспринимает устную речь, страдает произношение, так как нарушается восприятие собственной речи. Человек может говорить, излагать устно свои мысли, но не понимает чужой речи, и хотя слух и сохранен — человек не узнает слов. Такое вот состояние называется сенсорной слуховой афазией. Такой человек часто много говорит (логорея), но речь его неправильная (аграмматизм), при этом наблюдается замена слогов и слов (парафазии). Поле 37, отвечает за запоминание слов. Люди с поражениями этого поля не помнят названия предметов. При этом они очень напоминают забывчивых людей, которым постоянно приходится подсказывать нужные слова. Такой человек, забыв название предмета, четко помнит его назначение и свойства, поэтому долго описывает его качества, объясняет, что делают с этим предметом, но назвать его, хоть убей, не может. Ну, например, вместо слова «галстук» человек, глядя на него, говорит примерно следующее: «это то, что надевают на шею и завязывают специальным узлом, чтобы было красиво, когда идут в гости». Центр восприятия письменной речи – располагается в зрительной зоне коры головного мозга. На границе височной, теменной и затылочной долей (поле 39) находится центр чтения письменной речи, обеспечивающий распознавание и хранение образов письменной речи. Понятно, что поражения этого центра приводят к невозможности чтения и письма. При повреждении этого центра зрение сохранится, но тут же наступит расстройство узнавания – так называемая зрительная агнозия. Такой человек, будучи абсолютно грамотным, не сможет прочесть написанное, и будет в состоянии признать знакомого человека только после того, как тот заговорит. Таким образом, речевой процесс есть круговой процесс. Речевой круг образуют три мозговых речевых центра. центр Брока производит речь, управляя речевой мускулатурой, центр Вернике распознает собственную речь и речь других людей (слуховой центр речи), ассоциативный центр создает структуру фраз и предложений. 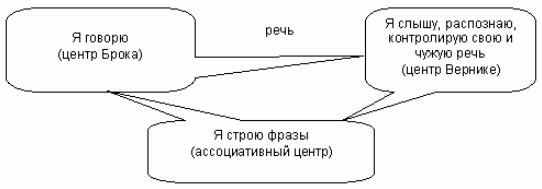 Разрыв речевого круга в любой точке разрушает речевой процесс. Примеры: 1. Глухота блокирует центр Вернике. Попытка восстановить речевой круг заставляет говорить громко. Абсолютная глухота делает человека немым (глухонемым) вследствие полного разрыва речевого круга на уровне центра Вернике. В неврологии данное состояние определяется как сенсорная афазия. 2. Центр Брока поражается при детском церебральном параличе. Тяжелая форма этого заболевания также резко нарушает или делает невозможным речевой процесс вследствие полного разрыва речевого круга на уровне центра Брока. В неврологии данное состояние определяется как моторная афазия. 3. Ассоциативный центр поражается при некоторых неврологических заболеваниях, травмах головного мозга. При этом нарушается способность составления фраз. Однако, данные нарушения наблюдаются не часто, т.к. ассоциативный центр менее жестко структурирован. 4. Заикание есть периодический разрыв речевого круга (не стабильная работа речевого круга). Цитоархитектонические поля по Бродману   Информационный минимум Лимбическая система мозга 1. Лимбическая система - функциональное объединение структур мозга, участвующих в организации эмоционально-мотивационного поведения, способствует научению и образованию следов долговременной памяти, обеспечивает общее усовершенствование приспособления организма к постоянно изменяющимся условиям окружающей среды. 2. Функции лимбической системы: · - мотивационно-эмоциональная; · - память и обучение; · - регуляция цикла «сон-бодрствование»; · -репродуктивное и сексуальное поведение. 3. Корковые структуры лимбической системы: гиппокамп, грушевидная доля коры, обонятельная луковица, поясная извилина, околопоясная извилина, орбитальная часть орбито-фронтальной коры, лобная и теменная доли коры. 4. Подкорковые стуктуры: гипоталамус, гиппокамп, миндалевидное тело, перегородка, передние ядра таламуса, ретикулярная формация среднего мозга, центральное серое вещество среднего мозга. 5. Особенность лимбической системы – наличие двухсторонних нейрональных связей между ее структурами ("кругов"), что дает возможность пролонгирования возбуждения за счет реверберации и повышения проводимости синапсов. 6. Большой круг Папеца: поясная извилина – гиппокамп - свод - мамиллярные телами гипоталамуса - передние ядра таламуса - поясная извилина. 7. Большой круг Папецаобеспечивает процессы запоминания и обучения. 8. Малый круг Наута: миндалевидное тело - гипоталамус - мезенцефальные структуры - миндалевидное тело) 9. Малый круг Наута регулирует агрессивно-оборонительные, пищевые и сексуальные формы поведения. 10. Большой круг Папеца и малый круг Наута связаны через гипоталамус. 11. Функции гиппокампа: обеспечение механизмов кратковременной и промежуточной памяти, обучения. 12. Разрушение гиппокампа - утрата способности к запоминанию при сохранении памяти о событиях, предшествовавших разрушению (антероградная амнезия). 13. Гиппокамп способен генерировать: · - высокоамплитудную ритмическую активность – быстрый бета-ритм (14-30 импульсов в 1 секунду) · - медленный тета-ритм (4-7 импульсов в 1 секунду). 14. Тета-ритм в гиппокампе отражает его реакцию: · - при формировании ориентировочного рефлекса, реакциях настороженности, повышении внимания и динамике обучения; · - при высоком уровне эмоционального напряжения – страхе, агрессии, голоде и жажде. 15. Гипоталамус обеспечивает вегетативные проявления эмоциональных реакций (покраснение кожи, потение и другие) 16. Гипоталамус имеет мотивационные центры (эмоция появляются, когда возникает мотивация, обусловленная потребностью). 17. Центры гипоталамуса обусловливают оценку ощущений «приятное» или «неприятное», удовлетворение и отвращение - это называют вознаграждением и наказанием. 18. Функции миндалевидного тела связаны с обеспечением оборонительного поведения, вегетативными, двигательными, эмоциональными реакциями, мотивацией условнорефлекторного поведения. 19. Повреждение миндалевидного тела приводит к исчезновению страха, успокоению, неспособности к ярости и агрессии, вызывает гиперсексуальность, гиперфагию. 20. Функциональная деление областей новой коры больших полушарий: сенсорные, моторные и ассоциативные. 21. Сенсорные области: · - соматосенсорная кора (задние центральные извилины) · - зрительная кора (затылочные доли) · - слуховая кора (часть височных долей). 22. Моторная кора находится в передних центральных извилинах и в примыкающих к ним спереди регионах лобных долей. 23. Ассоциативная кора: префронтальная кора лобных долей, теменно-височно-затылочная и лимбическая (внутренние и нижние поверхности лобных долей, внутренние поверхности затылочных долей и нижние отделы височных долей). 24. В сенсорной КБП 1 и 4 слои относительно широкие, принимающие таламокортикальные специфические и неспецифические проекции. 25. В моторной КБП начинаются кортикофугальные пирамидные и кортико-таламические пути, берущие начало в широких 5 и 6 слоях. 26. В ассоциативной КБП относительно широкие 2 и 3 слои КБП, клетки которых образуют обилие ассоциативных связей с соседними зонами. 27. Функционально в КБП выделяют 3 зоны: первичная, вторичная, третичная. 28. Первичная сенсорная КБП – это центральные отделы сенсорных систем, в них оканчиваются специфические таламокортикальные пути, имеют четкую соматотопическую организацию, выполняют первичный анализ сенсорной информации, нейроны мономодальны, моносенсорны. 29. Вторичная сенсорная КБП – зоны, расположенные вокруг первичных зон, выполняющие дальнейший анализ сенсорной информации, нейроны полимодальны, моносенсорны. 30. Третичная сенсорная зона КБП (ассоциативная КБП) – зона взаимного перекрытия сенсорных систем, осуществляет выработку условных рефлексов и интеллектуальной деятельности мозга, нейроны обладают высокой степенью конвергенции, полимодальны, полисенсорны. 31. Электроэнцефалография (ЭЭГ) - метод регистрации суммарной электрической активности различных отделов головного мозга, отражающий их функциональную активность, оценивается частота и амплитуда волн на ЭЭГ. 32. Ритмы ЭЭГ: · Альфа (α)-ритм: (8-13 Гц, 50-100 мкВ), выражен в затылочной и теменной областях коры и регистрируется в условиях физического и умственного покоя при закрытых глазах, при открытых глазах сменяется β-ритмом. Регистрируется у 85-95% здоровых взрослых. · Бета (β)-ритм: (14-50 Гц, до 25 мкВ) характерен для состояния интенсивной деятельности мозга при открытых глазах. Лучше всего β-ритм регистрируется в области центральных извилин, однако распространяется и на задние центральные и лобные извилины. β-Ритм связан с соматическими сенсорными и двигательными корковыми механизмами · Тета (θ)-ритм ( 4-8 Гц, 100-150 мкВ) регистрируется в височной и теменной областях при психомоторной активности, стрессе, во время засыпания, при гипоксии и лёгком наркозе. · Дельта(δ)-ритм ( 0,5-3,5 Гц, 250-300 мкВ) - глубокий сон, глубокий наркоз, гипоксия. 33. Реакция десинхронизации — соответствует изменению соответствующей ритмической активности ЭЭГ (засыпание, пробуждение, быстрый сон, реакция активации), проявляется изменением частоты ритма и амплитуды волн на ЭЭГ. 34. Речедвигательный центр Брока в нижней части лобных извилин (поле 44 левого полушария) — это двигательный центр мышц языка; при его поражении развивается моторная афазия (человек понимает речь, но сам говорить не может). 35. Сенсорный центр Вернике в верхней височной извилине (поля 22, 37, 42 левого полушария) обусловливает восприятие устной речи; при его поражении возникает сенсорная афазия (человек не воспринимает устную речь, страдает произношение). |