вопросы Вари. Вопросы Вари. По определению Штерн, гематоэнцефалический барьер
 Скачать 1.35 Mb. Скачать 1.35 Mb.
|

|
16.Гематоэнцефалический барьер: структура, функции, значение, результаты нарушения целостности. Глия: структура и функции. По определению Штерн, гематоэнцефалический барьер- это совокупность физиологических механизмов и соответствующих анатомических образований в центральной нервной системе, участвующих в регулировании состава цереброспинальной жидкости (ЦСЖ). Гематоэнцефалический барьер состоит из 5 слоев, из которых 3 образованы за счет трехслойной мембраны эндотелиоцита, а 2 других – двумя мембранными слоями астроцита. Морфологической основой ГЭБ является эндотелий капилляров, его базальная мембрана, прилегающие к ней отростки глиоцитов (сосудистые ножки) и перициты. По своей, ультраструктурной организации капилляры головного мозга относятся к гемокапиллярам с непрерывной эндотелиальной выстилкой и базальной мембраной. Эндотелий в морфологической структуре ГЭБ играет важную роль. Нормальные эндотелиальные клетки образуют высокоизбирательный барьер для прохождения в паренхиму мозга веществ, входящих в состав. Основу барьера составляют капилляры головного мозга, отличающиеся от капилляров других систем прочной межэндотелиальной связью, обусловленной отсутствием пор между эндотелиоцитами и сплошной базальной мембраной. Базальная мембрана эндотелиоцитов трехслойная, в ее дупликатуре располагаются перициты, содержащие мембранные органеллы и рибосомы. Для перицитов стенки капилляров мозга характерно наличие большого количества миофиломентов. Они участвуют в регуляции просвета микрососудов. К функции перицитов относится поддержание тонуса базальной мембраны, осуществление сократительной деятельности, а также опосредованное влияние на регенерацию эндотелия ГЭБ через секрецию трансформирующего фактора роста базальной мембраной и слоем астроцитарных отростков. Перициты участвуют в двигательной регуляции капилляров и имеют отношение к передаче им информации об изменении характера их метаболического окружения. На большей части поверхности сосудов располагаются отростки астроцитов, имеющие светлую цитоплазму, в которой выявляется умеренное количество митохондрий, канальцев эндоплазматической сети, везикулярные структуры, пиноцитозные везикулы, что указывает на участие астроцитов в транспорте веществ между капиллярами и нейронами Астроглия обеспечивает сохранение фенотипа ГЭБ и способствует регенерации его эндотелия. Таким образом, перициты, встроенные в базальную мембрану сосудов и переваскулярные клетки микроглии, а также астроциты играют главную роль в формировании плотных контактов ГЭБ. На современном этапе ведется изучение специфических трансмембранных белков (оклудин, клаудин) в эндотелиальных клетках головного мозга, участвующих в формировании твердых контактов, регулирующих проницаемость ГЭБ Основные функции гематоэнцефалического барьера: локальное управление кровотоком головного мозга, в соответствии с потребностями тех или иных анатомических областей; метаболическое обеспечение нейронов и метаболизма синапсов; защитная функция от нейротоксических веществ.  Гематоэнцефалический барьер ограничивает прохождение патогенов , диффузию растворенных веществ в крови и больших или гидрофильных молекул в спинномозговую жидкость , в то же время обеспечивая диффузию гидрофобных молекул (O 2 , CO 2 , гормоны) и малых неполярных молекул. молекулы. Клетки барьера активно транспортируют продукты метаболизма , такие как глюкоза, через барьер, используя специфические транспортные белки . Барьер также ограничивает прохождение периферических иммунных факторов, таких как сигнальные молекулы, антитела и иммунные клетки, в ЦНС, тем самым изолируя мозг от повреждений, вызванных периферическим иммунным процессом. 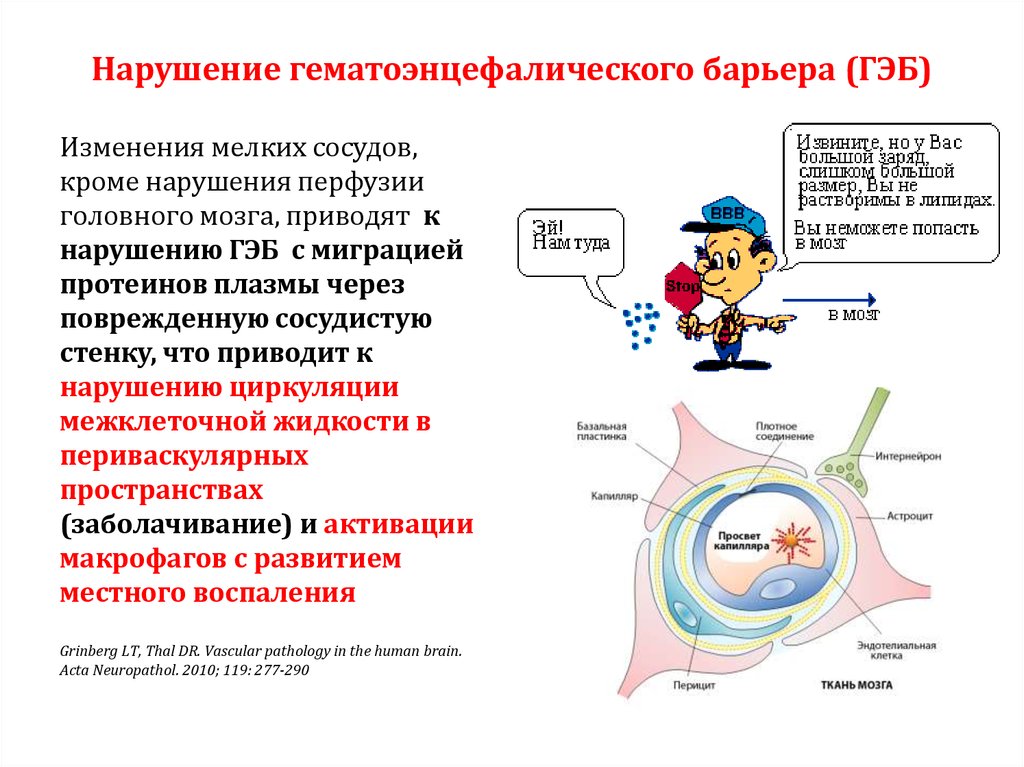 Н 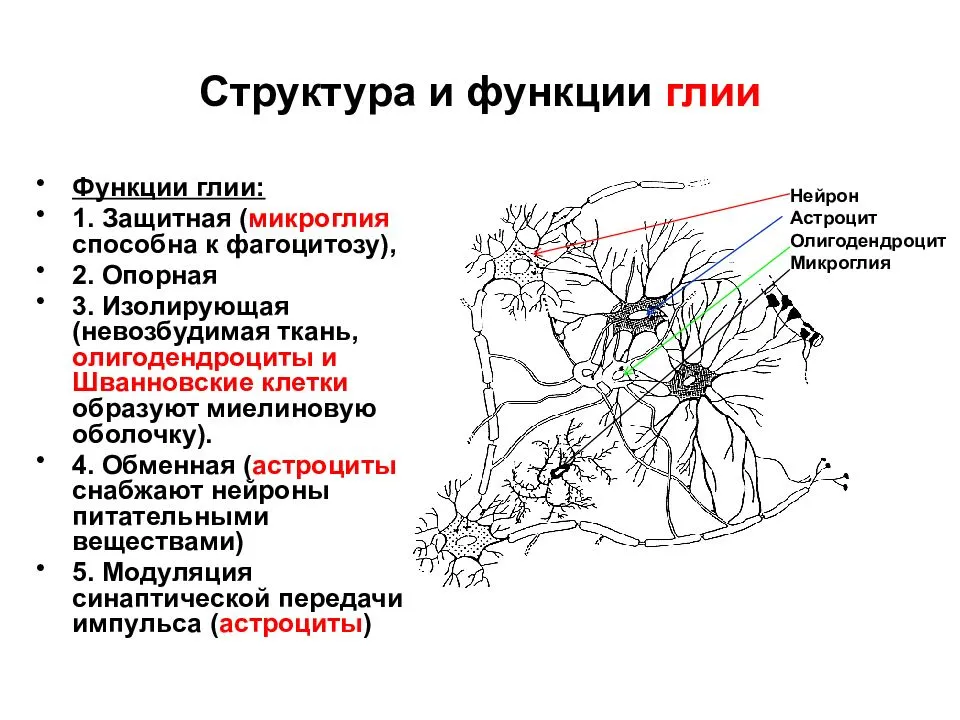 ЕЙРОГЛИ́Я (глия), клетки нервной ткани, выполняющие важнейшие функции по поддержанию процессов жизнедеятельности нейронов. Нейроглиальные клетки в 3–4 раза мельче нейронов, в отличие от последних обладают способностью к делению. Суммарно в мозге человека число клеток Н. (84,6 млрд.) и нейронов (86,1 млрд.) примерно одинаковое, но их соотношение в разных областях мозга меняется. ЕЙРОГЛИ́Я (глия), клетки нервной ткани, выполняющие важнейшие функции по поддержанию процессов жизнедеятельности нейронов. Нейроглиальные клетки в 3–4 раза мельче нейронов, в отличие от последних обладают способностью к делению. Суммарно в мозге человека число клеток Н. (84,6 млрд.) и нейронов (86,1 млрд.) примерно одинаковое, но их соотношение в разных областях мозга меняется.17. Нейрон как структурно-функциональная единица ЦНС. Виды нейронов. Интегративная функция нейрона. Структурной и функциональной единицей нервной системы является нервная клетка — нейрон. НЕЙРОНЫ — специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов. Уникальными особенностями нейрона являются способность генерировать электрические разряды и передавать информацию с помощью специализированных окончаний — синапсов. Функционально в нейроне выделяют следующие части: ВОСПРИНИМАЮЩУЮ — дендриты, мембрана сомы нейрона; ИНТЕГРАТИВНУЮ — сома с аксонным холмиком; ПЕРЕДАЮЩУЮ — аксонный холмик с аксоном. Сома обеспечивает также рост дендритов и аксона. Сома нейрона заключена в многослойную мембрану, обеспечивающую формирование и распространение электротонического потенциала к аксонному холмику. ВОСПРИНИМАЮЩАЯ ЧАСТЬ. ДЕНДРИТЫ — основное воспринимающее поле нейрона. Мембрана дендрита и синаптической части тела клетки способна реагировать на медиаторы, выделяемые аксонными окончаниями изменением электрического потенциала. Обычно нейрон имеет несколько ветвящихся дендритов. Необходимость такого ветвления обусловлена тем, что нейрон как ин формационная структура должен иметь большое количество входов. Информация к нему поступает от других нейронов через специализированные контакты, так называемые шипики. МЕМБРАНА СОМЫ НЕЙРОНА имеет толщину 6 нм и состоит из двух слоев липидных молекул. Гидрофильные концы этих молекул обращены в сторону водной фазы: один слой молекул обращен внутрь, другой – наружу. Гидрофильные концы повернуты друг к другу – внутрь мембраны. ИНТЕГРАТИВНАЯ ЧАСТЬ. АКСОНОВЫЙ ХОЛМИК – место выхода аксона из нейрона. СОМА НЕЙРОНА (тело нейрона) выполняет наряду с информационной и трофическую функцию относительно своих отростков и синапсов. Сома обеспечивает рост дендритов и аксонов. Сома нейрона заключена в многослойную мембрану, которая обеспечивает формирование и распространение электротонического потенциала к аксонному холмику. ПЕРЕДАЮЩАЯ ЧАСТЬ. АКСОН представляет собой вырост цитоплазмы, приспособленный для проведения информации, собранной дендритами, переработанной в нейроне и переданной аксону через аксонный холмик — место выхода аксона из нейрона. Аксон данной клетки имеет постоянный диаметр. Аксон имеет разветвленные окончания. АФФЕРЕНТНЫЕ НЕЙРОНЫ — нейроны, воспринимающие информацию. Как правило, афферентные нейроны имеют большую разветвленную сеть. Это характерно для всех уровней ЦНС. В задних рогах спинного мозга афферентными являются чувствительные нейроны малых размеров с большим числом дендритных отростков, в то время как в передних рогах спинного мозга эфферентные нейроны имеют тело большого размера, более грубые, менее ветвящиеся отростки. Эти различия нарастают по мере изменения уровня ЦНС к продолговатому, среднему, промежуточному, конечному мозгу. Наибольшие различия афферентных и эфферентных нейронов отмечаются в коре большого мозга. ВСТАВОЧНЫЕ НЕЙРОНЫ обрабатывают информацию, получаемую от афферентных нейронов, и передают ее на другие вставочные или на эфферентные нейроны. Вставочные нейроны могут быть возбуждающими или тормозными. Активация ВОЗБУЖДАЮЩИХ ВСТАВОЧНЫХ НЕЙРОНОВ в новой коре облегчает передачу информации с одной группы нейронов в другую. Причем это происходит за счет «медленных» пирамидных нейронов, способных к длительной тонической активации и поэтому передающих сигналы достаточно медленно и длительно. ТОРМОЗНЫЕ ВСТАВОЧНЫЕ НЕЙРОНЫ возбуждаются прямыми сигналами, идущими в их собственный центр, или сигналами, идущими из того же центра, но по обратным связям. Прямое возбуждение тормозящих вставочных нейронов характерно для промежуточных центров афферентных спиноцеребральных путей. ЭФФЕРЕНТНЫЕ НЕЙРОНЫ НЕРВНОЙ СИСТЕМЫ — это нейроны, передающие информацию от нервного центра к исполнительным органам или другим центрам нервной системы. Основной особенностью эфферентных нейронов является наличие длинного аксона, обладающего большой скоростью проведения возбуждения. Эфферентные нейроны разных отделов коры больших полушарий связывают между собой эти отделы по аркуатным связям. Нейроны автономной нервной системы, например ядер блуждающего нерва, боковых рогов спинного мозга, также относятся к эфферентным.   Интегративная функция- обобщение реакций нейрона на отдельные тормозные или возбуждающие влияния, в результате чего в ответной реакции нейрона (возбуждении или торможении его) «учитываются» все разнообразные сигналы, пришедшие к данному нейрону от других клеток. Интегративным процессам способствуют: а) суммация деполяризующих и гиперполяризующих токов на аксонном холмике, б) действие различных синаптических процессов на активацию (освобождение) одних и тех же внутриклеточных посредников: цАМФ, цГМФ, ионов кальция и др. 18. Синапсы в ЦНС: классификация и свойства. Возбуждающий постсинаптический потенциал, его свойства и значение. Возбуждающие медиаторы (нейротрансмиттеры) в ЦНС. Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку). Cтруктура синапса: 1) пресинаптическая мембрана (электрогенная мембрана в терминале аксона, образует синапс на мышечной клетке); 2) постсинаптическая мембрана (электрогенная мембрана иннервируемой клетки, на которой образован синапс); 3) синаптическая щель (пространство между пресинаптической и постсинаптической мембраной, заполнена жидкостью, которая по составу напоминает плазму крови). 2) Существует несколько классификаций синапсов. По локализации: 1) центральные синапсы;2) периферические синапсы. Центральные синапсы лежат в пределах центральной нервной системы, а также находятся в ганглиях вегетативной нервной системы. Центральные синапсы – это контакты между двумя нервными клетками, причем эти контакты неоднородны и в зависимости от того, на какой структуре первый нейрон образует синапс со вторым нейроном, различают: 1) аксосоматический, образованный аксоном одного нейрона и телом другого нейрона; 2) аксодендритный, образованный аксоном одного нейрона и дендритом другого; 3) аксоаксональный (аксон первого нейрона образует синапс на аксоне второго нейрона); 4) дендродентритный (дендрит первого нейрона образует синапс на дендрите второго нейрона). Различают несколько видов периферических синапсов: 1) мионевральный (нервно-мышечный), образованный аксоном мотонейрона и мышечной клеткой;2) нервно-эпителиальный, образованный аксоном нейрона и секреторной клеткой. Функциональная классификация синапсов:1) возбуждающие синапсы;2) тормозящие синапсы. По механизмам передачи возбуждения в синапсах:1) химические;2) электрические. Особенность химических синапсов заключается в том, что передача возбуждения осуществляется при помощи особой группы химических веществ – медиаторов. Различают несколько видов химических синапсов:1) холинэргические. В них происходит передача возбуждения при помощи ацетилхолина;2) адренэргические. В них происходит передача возбуждения при помощи трех катехоламинов;3) дофаминэргические. В них происходит передача возбуждения при помощи дофамина;4) гистаминэргические. В них происходит передача возбуждения при помощи гистамина;5) ГАМКэргические. В них происходит передача возбуждения при помощи гаммааминомасляной кислоты, т. е. развивается процесс торможения. Особенность электрических синапсов заключается в том, что передача возбуждения осуществляется при помощи электрического тока. Таких синапсов в организме обнаружено мало. Синапсы имеют ряд физиологических свойств: 1) клапанное свойство синапсов, т. е. способность передавать возбуждение только в одном направлении с пресинаптической мембраны на постсинаптическую; 2) свойство синаптической задержки, связанное с тем, что скорость передачи возбуждения снижается; 3) свойство потенциации (каждый последующий импульс будет проводиться с меньшей постсинаптической задержкой). Это связано с тем, что на пресинаптической и постсинаптической мембране остается медиатор от проведения предыдущего импульса; 4) низкая лабильность синапса (100–150 имульсов в секунду). Скорость проведения возбуждения через синапс намного меньше, чем по нервному волокну, так как здесь тратится время на активацию пресинаптической мембраны, переход через нее кальция, выделение ацетилхолина в синаптическую щель, деполяризацию постсинаптической мембраны, развитие ПКП. Синаптическая передача возбуждения имеет ряд свойств: 1) Наличие медиатора в пресинаптической части синапса; 2) Относительная медиаторная специфичность синапса, т. е. каждый синапс имеет свой доминирующий медиатор. 3) Переход постсинаптической мембраны под влиянием медиаторов в состояние де- или гиперполяризации; 4) Возможность действия специфических блокирующих агентов на рецептирующие структуры постсинаптической мембраны; 5) Увеличение длительности постсинаптического потенциала мембраны при подавлении действия ферментов, разрушающих синаптической медиатор; 6) Развитие в постсинаптической мембране ПСП из миниатюрных потенциалов, обусловленных квантами медиатора; 7) Зависимость длительности активной фазы действия медиатора в синапсе от свойств медиатора; 8) Односторонность проведения возбуждения; 9) Наличие хемочувствительных рецепторуправляемых каналов постсинаптической мембраны; 10) Увеличение выделения квантов медиатора в синаптическую щель пропорционально частоте приходящих по аксону импульсов; 11) Зависимость увеличения эффективности синаптической передачи от частоты использования синапса («эффект тренировки»); 12) Утомляемость синапса, развивающаяся в результате длительного высокочастотного его стимулирования. В этом случае утомление может быть обусловлено истощением и несвоевременным синтезом медиатора в пресинаптической части синапса или глубокой, стойкой деполяризацией постсинаптической мембраны (пессимальное торможение). 2) Перечисленные свойства относятся к химическим синапсам. Электрические синапсы имеют некоторые особенности, а именно: 1)малую задержку проведения возбуждения; 2)возникновение деполяризации как в пре-, так и в постсинаптической частях синапса; 3)наличие большей площади синаптической щели в электрическом синапсе, чем в химическом. Возбуждающий постсинаптический потенциал( ВПСП)В возбуждающих синапсах нервной системы медиатором может являться ацетилхолин, норадреналин, дофамин, серотонин, глугаминовая кислота, вещество Р, а также большая группа других веществ, являющихся, если не медиаторами в прямом значении, то во всяком случае модуляторами (меняющими эффектиьность) синаптической передачи. Возбуждающие медиаторы вызывают появление на постсинаптической мембраневозбуждающего постсинаптического потенциала (ВПСП). Его формирование обусловлено тем, что медиатор-рецепторный комплекс активирует Na- каналы мембраны (а также вероятно и Са-каналы) и вызывает за счет поступления натрия внутрь клетки деполяризацию мембраны. Одновременно происходит и уменьшение выхода из клетки ионовК+Амплитуда одиночного ВПСП однако довольно мала, и для уменьшения заряда мембраны до критического уровня деполяризации необходима одновременная активация нескольких возбуждающих синапсов.ВПСП, образуемые на постсинаптической мембране этих синапсов, способны суммироваться, т.е. усиливать друг друга, приводя к росту амплитуды ВПСП (пространственная суммация).Растет амплитуда ВПСП и при увеличении частоты поступающих к синапсу нервных импульсов (временная суммация), что повышает число выводимых в синаптическую щель квантов медиатора.К возбуждающим нейромедиаторам относят ацетилхолин, дофамин, серотонин, глутаминовую кислоту, норадреналин. В некоторых синапсах в качестве возбуждающего нейромедиатора могут выделяться пурины, АТФ, некоторые нейропептиды. К тормозным нейромедиаторам относят гамма-аминомасляную кислоту (ГАМК) и глицин. 19. Торможение в ЦНС: виды и механизмы. Роль И.М. Сеченова. Тормозной постсинаптический потенциал, его свойства, значение. Торможение – активный процесс, возникающий при действии раздражителей на ткань, проявляется в подавлении другого возбуждения, функционального отправления ткани нет. Торможение может развиваться только в форме локального ответа. Выделяют два типа торможения: 1) первичное. Для его возникновения необходимо наличие специальных тормозных нейронов. Торможение возникает первично без предшествующего возбуждения под воздействием тормозного медиатора. Различают два вида первичного торможения: · пресинаптическое в аксо-аксональном синапсе; · постсинаптическое в аксодендрическом синапсе. 2) вторичное. Не требует специальных тормозных структур, возникает в результате изменения функциональной активности обычных возбудимых структур, всегда связано с процессом возбуждения. Виды вторичного торможения: · запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за пределами работоспособности нейрона; · пессимальное, возникающее при высокой частоте раздражения; парабиотическое, возникающее при сильно и длительно действующем раздражении; · торможение вслед за возбуждением, возникающее вследствие снижения функционального состояния нейронов после возбуждения; · торможение по принципу отрицательной индукции; · торможение условных рефлексов. Процессы возбуждения и торможения тесно связаны между собой, протекают одновременно и являются различными проявлениями единого процесса. Очаги возбуждения и торможения подвижны, охватывают большие или меньшие области нейронных популяций и могут быть более или менее выраженными. Возбуждение непременно сменяется торможением, и наоборот, т. е. между торможением и возбуждением существуют индукционные отношения. Торможение лежит в основе координации движений, обеспечивает защиту центральных нейронов от перевозбуждения. Торможение в ЦНС может возникать при одновременном поступлении в спинной мозг нервных импульсов различной силы с нескольких раздражителей. Более сильное раздражение тормозит рефлексы, которые должны были наступать в ответ на более слабые. В 1862 г. И. М. Сеченов открыл явление центрального торможения. Он доказал в своем опыте, что раздражение кристалликом хлорида натрия зрительных бугров лягушки (большие полушария головного мозга удалены) вызывает торможение рефлексов спинного мозга. После устранения раздражителя рефлекторная деятельность спинного мозга восстанавливалась. Результат этого опыта позволил И. М. Сеченому сделать заключение, что в ЦНС наряду с процессом возбуждения развивается процесс торможения, который способен угнетать рефлекторные акты организма. Н. Е. Введенский высказал предположение, что в основе явления торможения лежит принцип отрицательной индукции: более возбудимый участок в ЦНС тормозит активность менее возбудимых участков. Современная трактовка опыта И. М. Сеченова (И. М. Сеченов раздражал ретикулярную формацию ствола мозга): возбуждение ретикулярной формации повышает активность тормозных нейронов спинного мозга – клеток Реншоу, что приводит к торможению α-мотонейронов спинного мозга и угнетает рефлекторную деятельность спинного мозга. Тормозные синапсы образованы специальными тормозными нейронами (точнее, их аксонами). Медиатором могут быть глицин, ГАМК и ряд других веществ. Обычно глицин вырабатывается в синапсах, с помощью которых осуществляется постсинаптическое торможение. При взаимодействии глицина как медиатора с глициновыми рецепторами нейрона возникает гиперполяризация нейрона (ТПСП) и, как следствие, - снижение возбудимости нейрона вплоть до полной его рефрактерности. В результате этого возбуждающие воздействия, оказываемые через другие аксоны, становятся малоэффективными или неэффективными. Нейрон выключается из работы полностью. Тормозные синапсы открывают в основном хлорные каналы, что позволяет ионам хлора легко проходить через мембрану. Чтобы понять, как тормозные синапсы тормозят постсинаптический нейрон, нужно вспомнить, что мы знаем о потенциале Нернста для ионов Сl-. Мы рассчитали, что он равен примерно -70 мВ. Этот потенциал отрицательнее, чем мембранный потенциал покоя нейрона, равный -65 мВ. Следовательно, открытие хлорных каналов будет способствовать движению отрицательно заряженных ионов Сl- из внеклеточной жидкости внутрь. Это сдвигает мембранный потенциал в направлении более отрицательных значений по сравнению с покоем приблизительно до уровня -70 мВ. Открытие калиевых каналов позволяет положительно заряженным ионам К+ двигаться наружу, что приводит к большей отрицательности внутри клетки, чем в покое. Таким образом, оба события (вход ионов Сl- в клетку и выход ионов К+ из нее) увеличивают степень внутриклеточной отрицательности. Этот процесс называют гиперполяризацией. Увеличение отрицательности мембранного потенциала по сравнению с его внутриклеточным уровнем в покое тормозит нейрон, поэтому выход значений отрицательности за пределы исходного мембранного потенциала покоя называют ТПСП. Тормозной постсинаптический потенциал. Тормозной постсинаптический потенциал - гиперполяризация постсинаптической мембраны втормозных синапсах . По временному ходу тормозной постсинаптический потенциал представляет собой зеркальное отображение возбуждающего постсинаптического потенциала со временем нарастания и спада соответственно 1-2 и 10-12 мс. Сдвиг проводимостипостсинаптической мембраны длится также около 1-2 мс. +При действии тормозных нейромедиаторов в постсинаптической мембране открываются каналы для ионов хлора, вследствие чего ионы хлора входят в клетку, отрицательный заряд на внутренней стороне мембраны увеличивается и происходит гиперполяризация мембраны – образуется тормозной постсинаптический потенциал (ТПСП), который затрудняет образование ПД. 20. Свойства нервных центров: суммация, трансформация ритма, дивергенция, конвергенция, иррадиация и др. Доминанта, значение работ А.А. Ухтомского. Нервным центром называется функциональное объединение нейронов, обеспечивающее осуществление какого-либо рефлекса или регуляцию какой-либо определенной функции. Нейроны, входящие в нервный центр, обычно находятся в одном отделе ЦНС, но могут располагаться и в нескольких. Центр дыхания располагается в средней трети продолговатого мозга, центр мочеиспускания — в крестцовом, центр коленного рефлекса — в поясничном отделе спинного мозга. В осуществлении сложных рефлексов целостного организма принимают участие, как правило, не один, а многие центры, расположенные в разных отделах мозга, включая его высшие отделы. Например, в акте дыхания участвует не только центр дыхания в продолговатом мозге, но и нервные клетки варолиева моста, коры головного мозга и мотонейроны спинного мозга. Особенности распространения возбуждения в ЦНС в основном определяются свойствами нервных центров: 1.Одностороннее проведение возбуждения. В ЦНС возбуждение может распространяться только в одном направлении: от рецепторного нейрона через вставочный к эфферентному нейрону, что обусловлено наличием синапсов. 2.Более медленное проведение возбуждения по сравнению с нервными волокнами. Промежуток времени от момента нанесения раздражения на рецептор до ответной реакции исполнительного органа называется временем рефлекса. Большая его часть тратится на проведение возбуждения в нервных центрах, где возбуждение проходит через синапсы. На выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5-2 мс (синаптическая задержка). Чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса. 3 Суммация возбуждений (или торможения). Нервные центры могут суммировать афферентные импульсы, что проявляется в усилении рефлекса при увеличении частоты раздражений или числа раздражаемых рецепторов. Различают два вида суммации: временная суммация — если импульсы приходят к нейрону по одному и тому же пути через один синапс с коротким интервалом, происходит суммирование ВПСП на постсинаптической мембране и она деполяризуется до уровня, достаточного для генерации ПД пространственная суммация связана с суммированием ВПСП, возникающих одновременно в разных синапсах одного нейрона. Оба вида суммации происходят в области аксонного холмика, где и генерируется ПД. 4. Конвергенция. В нервном центре несколько клеток могут передавать импульсы к одному нейрону, т. е. возбуждения конвергируют на нем. Конвергенция может быть результатом прихода возбуждающих или тормозных входных сигналов от различных источников. Так, моторные нейроны спинного мозга могут получать импульсы: от периферических нервных волокон, входящих в спинной мозг; волокон, соединяющих сегменты спинного мозга; кортикоспинальных волокон от коры мозга; тормозных путей от ретикулярной формации. В результате конвергенции происходит суммация импульсов от этих источников и возникает ответ, являющийся суммарным эффектом разнородной информации. 5. Дивергенция и иррадиация. Возбуждение даже единственного нервного волокна, по которому импульсы поступают в нервный центр, может послужить причиной возбуждения множества выходящих из центра нервных волокон. Морфологическим субстратом широкого распространения импульсов (иррадиации) возбуждения является ветвление аксонов и наличие большого числа вставочных нейронов в пределах центра. 6. Облегчение и окклюзия. На нейронах нервных центров оканчиваются не только волокна их собственных афферентных входов. Каждый из них получает веточки от афферентов соседнего центра, что может обусловливать развитие окклюзии (закупорки) или, наоборот, облегчения. Феномен окклюзии состоит в том, что количество возбужденных нейронов при одновременном раздражении афферентных входов обоих нервных центров оказывается меньше, чем арифметическая сумма возбужденных нейронов при раздельном раздражении каждого афферентного входа в отдельности. Явление окклюзии приводит к снижению силы суммарной ответной реакции. Феномен центрального облегчения характеризуется противоположным эффектом. На облегчении основано проторение пути - распространение возбуждения не по той цепи нейронов, по которой информация не поступала ранее, а через нейроны, уже облегченные первым раздражением. 7.Трансформация ритма возбуждений. Если сопоставить частоту импульсов в задних (чувствительных) и передних (двигательных) корешках спинного мозга при раздражении рецепторов, обычно их ритм не совпадает. Центры способны как снижать, таки повышать ритмы возбуждений, поступающих от рецепторов. 8.Рефлекторное последействие. Продолжительность рефлекса всегда больше, чем время раздражения, так как возбуждение нервных центрах сохраняется в течение некоторого времени после прекращения действия раздражителя. Это объясняется тем, чтовставочные нейроны в центре образуют замкнутые цепи («нейронные ловушки»), по которым ПД могут длительно циркулировать. 9.Высокая чувствительность к недостатку кислорода. Уменьшение доставки к клеткам мозга кислорода быстро ведет к тяжелым расстройствам деятельности ЦНС и гибели нейроном Кратковременное нарушение кровоснабжения мозга вследствие временного спазма его сосудов или падения давления крови приводит к потере сознания — обмороку. Своевременно приняты меры по восстановлению кровоснабжения мозга (нашатырный спирт, кофеин, горизонтальное положение тела и др.) выводя больного из обморока. 10.Высокая чувствительность к химическим веществам объясняется большим числом синапсов. На одном нейроне могут располагаться синапсы, обладающие чувствительностью к различным химическим веществам. Подбирая фармакологические препараты, которые избирательно блокируют одни синапсы, оставляядругие в рабочем состоянии, можно корректировать реакции организма. 11.Низкая функциональная подвижность (лабильность) и высокая утомляемость. Нервные центры, как и синапсы, обладают низкой функциональной подвижностью и быстрой утомляемостью в отличие от нервных волокон, которые считаются практически неутомляемыми и имеют высокую лабильность. 12.Посттетаническая потенциация — явление усиления рефлекторного ответа после длительного ритмического раздражения нервного центра. Это связано с сохранением ВПСП на нейронах центра в течение некоторого времени, что облегчает проведение последующих возбуждений через синапсы. 13. Тонус нервных центров. Регистрация биоэлектрической активности ЦНС даже при отсутствии раздражений показывает, что многие нервные центры (составляющие их нейроны) постоянно генерируют импульсы. Эта импульсация поступает к рабочим органам и свидетельствует о существовании некоторого постоянного тонического возбуждения нервных центров. 14. Пластичность. Нервные центры обладают способностью изменять собственное функциональное назначение и расширять свои функциональные возможности, т.е. существенно модифицировать картину осуществляемых рефлекторных реакций. Пластичность нервных центров тесно связана с изменением эффективности или направленности связей между нейронами. Особое внимание должен привлечь феномен доминанты, сформулированный академиком А. А. Ухтомским как временно господствующая рефлекторная система, направляющая работу нервных центров в данный момент. Интимные механизмы доминанты не определены, феноменология же ее вполне конкретна. Предварительно подготовленное импульсами своей рефлекторной дуги, подкрепленное в результате суммации влияниями из постороннего источника, возникающее в доминантном очаге возбуждение тормозит прочие рефлексы на очередные раздражители. При объединении нескольких локальных сетей формируются нервные центры, которые представляют собой комплекс элементов, необходимых и достаточных для осуществления определенного рефлекса или более сложного поведенческого акта. В свою очередь, нервные центры, расположенные в различных отделах мозга, могут кооперироваться в так называемые распределенные системы, которые координируют деятельность организма в целом. Эти системы имеют иерархическую структуру и представляют собой следующий, более высокий интегративный уровень ЦНС, деятельность которого базируется на некоторых общих принципах работы нервных центров. Одним из таких принципов является открытый А. А. Ухтомским принцип доминанты. Под доминантой А. А. Ухтомский понимал господствующий очаг возбуждения, предопределяющий характер текущих реакций центров в данный момент. Такой господствующий, или доминантный, центр может возникнуть в различных этажах ЦНС при достаточно длительном действии гуморальных или рефлекторных раздражителей и характеризуется: 1) наличием повышенной возбудимости; 2) инерционностью, обусловленной длительными следовыми процессами; 3) способностью к суммации и сопряженному торможению других центров, функционально несовместимых с деятельностью центров доминантного очага. В целом доминанта как состояние характеризуется своей направленностью и создает определенный вектор поведения. Рассматривая доминанту как общий принцип работы нервных центров, А. А. Ухтомский считал, что она связана с возбуждением целого созвездия, или констелляции нервных центров, которые временно кооперируются при выполнении биологически важной функции. Это созвездие создает динамический функциональный орган, сообщающий организму единство действия в данный момент. Формирование констелляции нервных центров, согласно А. А. Ухтомскому, может происходить за счет импульсных взаимодействий и сонастраивания работающих центров на единый ритм активности (усвоение ритма). 21. Центральное торможение. Тормозные медиаторы ЦНС, механизмы их действия. Вторичное торможение, его виды и физиологическое значение. Впервые идею о том, что в ЦНС помимо процессов возбуждения существует процесс торможения, высказал И.М. Сеченов. Если рассмотреть «архитектуру» использования тормозных нейронов при организации нейронных сетей, цепей и рефлекторных дуг, то можно выделить ряд вариантов этой организации (это отражается в названии данного вида торможения). 1. Реципрокное торможение. Как пример: сигнал от мышечного веретена поступает с афферентного нейрона в спинной мозг, где переключается на альфа-мотонейрон сгибателя и одновременно на тормозной нейрон, который тормозит активность альфа-мотонейрона разгибателя. Явление открыто Ч. Шеррингтоном. 2. Возвратное торможение. Альфа-мотонейрон, к примеру, посылает аксон к соответствующим мышечным волокнам. По пути от аксона отходит коллатераль, которая возвращается в ЦНС - она заканчивается на тормозном нейроне (клетка Реншоу) и активирует ее. Тормозной нейрон вызывает торможение альфа-мотонейрона, который запустил всю эту цепочку. Таким образом, альфа-мотонейрон, активируясь, через систему тормозного нейрона сам себя тормозит. 3. Существует ряд вариантов возвратного торможения, в частности, латеральное торможение. Суть его сводится к тому, что активируется, например, фоторецептор, он активизирует биполярную клетку, и одновременно активируется рядом расположенный тормозной нейрон, который блокирует проведение возбуждения от соседнего фоторецептора к ганглиозной клетке. Этим самым происходит «вытормаживание» информации в соседних участках. Таким способом создаются условия для четкого видения предмета (две точки на сетчатке рассматриваются как две раздельные точки в том случае, если между ними есть невозбужденные участки).  Вторичное торможение возникает в обычных возбудимых структурах и связано с процессом возбуждения. Виды вторичного поражения. Запредельное торможение - возникает в нейронах центральной нервной системы в том случае, когда поток информации к телу нейрона выше его работоспособности. Развивается резкое снижение возбудимости нейрона. Парабиотическое торможение - возникает при действии сильных и длительно действующих раздражителей (парабиоз в тканях). Парабиоз - явление пограничного состояния между гибелью и жизнью ткани (резко падают все свойства ткани, основное свойство - фазное изменение лабильности). Если парабиотический фактор продолжает действовать, ткань гибнет. Пессимальное торможение - возникает в синапсах центральной нервной системы при действии сильных и частых раздражителей. Торможение вслед за возбуждением - угнетение нейронов после возбуждения. Результат того, что вслед за пиком потенциала действия возникает период следовой гиперполяризации, который характеризуется снижением возбудимости. Торможение по принципу отрицательной индукции. Условное (внутреннее) торможение. 23. Спинальные рефлексы: классификация, механизмы контроля мышечного тонуса и фазных движений. Спинальный шок, механизм его развития. Проприорецепторы скелетных мышц и их роль в координации рефлексов. Виды спинальных рефлексов. Ч. Шеррингтон (1906) установил основные закономерности его рефлекторной деятельности и выделил основные виды осуществляемых им рефлексов. Собственно рефлексы мышц (тонические рефлексы) возникают при раздражении рецепторов растяжения мышечных волокон и сухожильных рецепторов. Они проявляются в длительном напряжении мышц при их растяжении. Защитные рефлексы представлены большой группой сгибательных рефлексов, предохраняющих организм от повреждающего действия чрезмерно сильных и опасных для жизни раздражителей. Ритмические рефлексы проявляются в правильном чередовании противоположных движений (сгибание и разгибание), сочетающихся с тоническим сокращением определенных групп мышц (двигательные реакции чесания и шагания). Рефлексы положения (позные) направлены на длительное поддержание сокращения групп мышц, придающих телу позу и положение в пространстве. Следствием поперечной перерезки между продолговатым и спинным мозгом является спинальный шок. Он проявляется резким падением возбудимости и угнетением рефлекторных функций всех нервных центров, расположенных ниже места перерезки Миостатический рефлекс осуществляется при функциональных состояниях, связанных с растяжением жевательной мускулатуры, начало ему дают импульсы от проприорецепторов жевательных мышц и их сухожилий. В деятельности жевательной системы имеет место сочетание различных рефлексов. Особое внимание заслуживает совокупность рефлексов, связанных с разобщением прикуса,10 которое играет важную роль в клинике стоматологии. Рефлексы разобщения прикуса являются постоянным спутником при всевозможных актах, связанных с длительным опусканием нижней челюсти и расположением ее на расстоянии больше 4-6 мм от верхней челюсти. Такое статическое состояние опущенное нижней челюсти, превышающее исходное положение при физиологическом покое, ведет к проявлению тонических рефлексов, возникающих с различных рецептивных полей жевательной системы (мышц, сухожилий, периодонта, слизистой рта). В формировании «рефлексов разобщения прикуса» большое участие принимают миостатические рефлексы жевательной мускулатуры - т.н. «рефлексы на растяжение». Характер проявления «рефлексов разобщения прикуса» зависит от степени опускания нижней челюсти, сочетания с другими рефлексами (периодонто-мускулярным, гингиво-мускулярным), чередования периодов покоя и раздражения, адаптации рецепторов к протезам и от индивидуальных особенностей больных |