репликация днк. доклад репликация днк. Репликация днк. Хроматин
 Скачать 152.33 Kb. Скачать 152.33 Kb.
|

|
Негосударственное образовательное учреждение высшего профессионального образования «Самарский медицинский институт «РЕАВИЗ» ДОКЛАД по дисциплине: биология на тему: «Репликация ДНК. Хроматин» Студентка: Сидоренко Екатерина Владимировна Факультет: Стоматологический Группа: 21-2003 Курс 1 Самара 2021г Репликация ДНК – это молекулярный процесс точного копирования молекул ДНК (ее нуклеотидной последовательности). С помощью механизма репликации происходит точная передача генетической информации от клетки к клетке и, таким образом, все клетки многоклеточного организма являются носителями одной и той же наследственной информации. Процесс синтеза ДНК сопровождается множеством событий и является, как правило, точным. Из одной молекулы ДНК синтезируются две идентичные дочерние молекулы. Этот процесс становится возможным благодаря структурным особенностям молекулы ДНК: - двухцепочечная структура; - комплементарность и антипараллельность. Основными характеристиками репликации являются: синтез ДНК является полуконсервативным, так как каждая цепь служит матрицей для синтеза дочерней цепи; репликация носит двунаправленный характер; синтез новой цепи осуществляется только в направлении 5'→3'; в репликации участвует значительное количество белковых факторов. Белки аппарата репликации В репликации ДНК участвует большое количество белковых факторов и ферментов ДНК-геликазы осуществляют деспирализацию и денатурацию участка ДНК с использованием энергии гидролиза АТФ и образование репликационной вилки. В связи с наличием двух репликационных вилок, существуют две геликазы, которые передвигаются в разных направлениях от точки инициации репликации. Праймаза – фермент, который инициирует синтез ДНК за счет синтеза небольшого фрагмента, состоящего из 11-12 рибонуклеотидов, - праймера или затравки (РНК-праймер). Геликаза совместно с праймазой образует комплекс - праймосома. Топоизомераза I разрывает только одну из двух це-пей двойной спирали ДНК, расщепляя фосфодиэфирные связи, что предупреждает суперспирализацию молекулы ДНК. Топоизомераза II ковалентно связывается с обеими цепями двойной спирали ДНК и вносит в нее на время двухцепочечный разрыв.  Рис. 7.2. Стабилизация одноцепочечной молекулы ДНК в процессе репликации Белки, связывающиеся с одноцепочечной ДНК (SSB – белки), это факторы, стабилизирующие одноцепочную ДНК в области денатурации и препятствующие комплементарному спариванию двух цепей или различных участков одной и той же цепи, имеющих палиндромные последовательности ДНК-полимеразы - ферменты, способные синтези-ровать новые цепи ДНК с матричной цепи. Полимеразная активность характеризуется следующими особенностями: синтез осуществляется только в направлении 5'→3' с присоединением нуклеотидов к группе 3'-ОН пентозы (рис.7.3); чтение матрицы происходит только в направлении 3'→5'; для синтеза используются предшественники трифосфатов, которые теряют во время реакции две фосфатные группы; ДНК-полимераза не может начать синтез новой полинуклеотидной цепи, а способна только добавлять новые нуклеотиды к существующей цепи или к РНК-праймеру. Наряду с синтетической активностью, ДНК-полимераза содержит субъединицы, которые обладают нуклеазной активностью для удаления РНК-затравок или нуклеотидов в процессе коррекции ошибок репликации ДНК. ДНК-лигаза – это фермент, связывающий участки вновь синтезированной ДНК, посредством образования фосфодиэфирных связей 3'→5'. 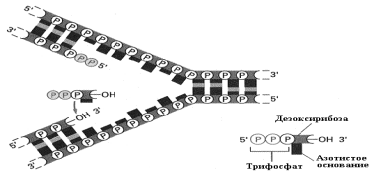 Механизм репликации Синтез начинается с момента деспирализации цепей ДНК и образования репликационной вилки. Каждая цепь представляет матрицу для вновь синтезируемой цепи. Деспирализация двух цепей ДНК необходима для того, чтобы каждое основание обеих цепей оказалось доступным для узнавания новыми основаниями и комплементарного спаривания. Синтез осуществляется в двух направлениях; от каждой точки инициации образуются две репликационные вилки в противоположном направлении. Для инициации репликации необходим белковый комплекс, названный реплисомой, который узнает точку инициации и запускает механизм репликации. Белок / белки, распознающие точку инициации (у дрожжей известно 5 белков), связываются с ori (Точка начала репликации, точка ori.)и инициируют деспирализацию ДНК в сайте DUE (DUЕ – участок ДНК, который легко деспирализуется, чувствителен к нуклеазам и не чувствителен к мутациям). Реплисома движется вдоль ДНК и осуществляет синтез обеих цепей репликационной вилки. Репликация представляет собой непрерывный рост обеих цепей спирали ДНК. Необходимо подчеркнуть что: - считывание информации с матрицы осуществляется только в направлении 3'→5'; - синтез новой цепи осуществляется только в направлении 5'→3'. Репликационная вилка асимметрична (рис.7.4). Из двух синтезированных дочерних цепей ДНК:  - одна цепь синтезируется непрерывно в направлении 5'→3' и называется ведущей (лидирующей) цепью. Она считывается в направлении 3'→5' с матричной цепи 3'→5'; - другая цепь синтезируется с матричной цепи 5'→3'. Считывание происходит также в направлении 3'→5', а новую цепь называют отстающей цепью. Она синтезируется прерывисто, с образованием фрагментов Оказаки. Длина фрагментов Оказаки у прокариот составляет - 1000-2000, а у эукариот – 100-200 нуклеотидов. Согласно гипотезе Артура Корнберга отстающая цепь развернута на 1800. Таким образом, молекулы ДНК-полимераз совместно с другими белками реплисомы обеспечивают одновременный синтез обеих цепей (рис. 7.5). 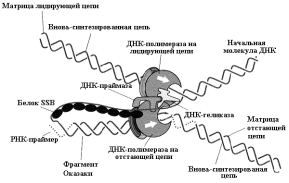 При встрече двух репликационных вилок процесс репликации останавливается, фермент ДНК-лигаза связывает все фрагменты, предварительно удаляются праймеры и заполняются недостающие фрагменты. Этапы репликации 1. Инициация включает следующие процессы: присоединение реплисомы к точке инициации репликации и деспирализация участка двойной спирали ДНК геликазами; синтез РНК-праймеров ферментом праймазой, выполняющей функции РНК-полимеразы; присоединение дезоксирибонуклеотидов, комплементарных матрице, к 3'-концу праймера ДНК-полимеразой. 2. Элонгация характеризуется удлинением вновь синтезируемых цепей, которое осуществляется ДНК-полимеразой, входящей в состав реплисомы, во время ее продвижения по матрице: непрерывный рост лидирующей цепи; прерывистый синтез фрагментов Оказаки на отстающей цепи; контроль ошибок спаривания оснований во время ре-пликации и их исправление при помощи 3'→5' экзо-нуклеазы из состава ДНК-полимеразы. 3. Терминация включает следующие процессы: удаление РНК-праймеров 5'→3' экзонуклеазой, представляющей собой субъединицу ДНК-полимеразы; заполнение недостающих участков ДНК-полимеразой; связывание фрагментов вновь синтезированной ДНК с помощью лигаз. Хроматин Хроматин представляет собой массу генетического вещества, состоящего из ДНК и белков, которые конденсируются с образованием хромосом во время деления эукариотических клеток. Хроматин содержится в ядре наших клеток. Основная функция хроматина состоит в том, чтобы сжать ДНК в компактную единицу, которая будет менее объемной и сможет войти в ядро. Хроматин состоит из комплексов небольших белков, известных как гистоны и ДНК. Гистоны помогают организовать ДНК в структуры, называемые нуклеосомами, обеспечивая фундамент для обертывания ДНК. Нуклеосома состоит из последовательности нитей ДНК, которые обертываются вокруг набора из восьми гистонов, называемых октомерами. Нуклеосома дополнительно складывается с получением хроматинового волокна. Хроматиновые волокна свертываются и конденсируются с образованием хромосом. Хроматин позволяет осуществить ряд клеточных процессов, включая репликацию ДНК, транскрипцию, восстановление ДНК, генетическую рекомбинацию и деление клеток. Эухроматин и гетерохроматин Хроматин внутри клетки может быть уплотнен в различной степени в зависимости от стадии клетки в клеточном цикле. Хроматин в ядре содержится в виде эухроматина или гетерохроматина. Во время интерфазы, клетка не делится, а подвергается периоду роста. Большая часть хроматина находится в менее компактной форме, известной как эухроматин. ДНК подвергается воздействию эухроматина, что позволяет проводить репликацию и транскрипцию ДНК. Во время транскрипции двойная спираль ДНК разматывается и открывается, чтобы можно было скопировать гены, кодирующие белки. Репликация и транскрипция ДНК необходимы для того, чтобы клетка синтезировала ДНК, белки и органеллы при подготовке к делению клеток (митоз или мейоз). Небольшой процент хроматина существует как гетерохроматин во время интерфазы. Этот хроматин плотно упакован, что не позволяет проводить транскрипцию гена. Гетерохроматин окрашивается красителями в более темный цвет, чем эухроматин. Хроматин в митозе: Профаза Во время профазы митоза волокна хроматина превращаются в хромосомы. Каждая реплицированная хромосома состоит из двух хроматид, соединенных в центромере. Метафаза Во время метафазы хроматин становится чрезвычайно сжатым. Хромосомы выровнены на метафазной пластинке. Анафаза Во время анафазы парные хромосомы (сестринские хроматиды) отделяются и вытягиваются микротрубочками веретена деления на противоположные полюса клетки. Телофаза В телофазе каждая новая дочерняя хромосома перемещается в свое собственное ядро. Хроматиновые волокна разматываются и становятся менее уплотненными. После цитокинеза образуются две генетически идентичные дочерние клетки. Каждая клетка имеет одинаковое количество хромосом. Хромосомы продолжают разматывать и удлинять образующий хроматин. |