Справочный материал. Глава 05 – Физиология нейронов. Справочный материал по Физиологии. Глава 5 Физиология нейронов
 Скачать 214.5 Kb. Скачать 214.5 Kb.
|

|
Справочный материал по Физиологии. Глава 5 – Физиология нейронов. Нервная система состоит из двух типов клеток — нервных (нейроны) и глиальных (нейроглия). К нейроглии относятся астроциты, микроглия, миелинобразующие клетки (олигодендроциты ЦНС и шванновские клетки периферических нервов). По отношению к нейронам глиоциты выполняют трофическую, опорную и изолирующую (электрический изолятор) функции. Нейрон — основная структурно-функциональная единица нервной системы. Число нейронов в мозге человека превышает 100109. Основная функция нейронов — генерация, передача и интеграция нервных импульсов. Нейроны В нервной клетке различают тело (перикарион) и отростки — дендриты и аксон (рис. 5–1). Как правило, нейрон имеет несколько дендритов и один аксон. Отростки нейрона и поверхность перикариона участвуют в образовании синапсов. Нейроны — возбудимые клетки — осуществляют передачу электрических сигналов в пределах самой нервной клетки (а между нейронами при помощи нейромедиаторов в синапсах) и этим обеспечивают способность мозга к переработке информации. 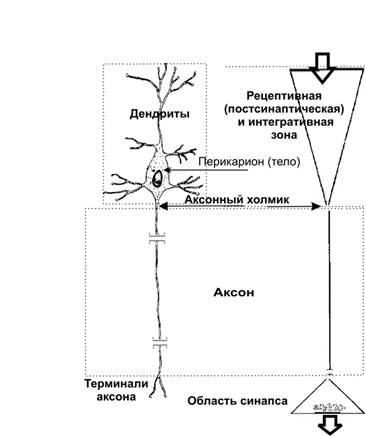 Рис. 5–1. Многоотростчатый нейрон [11]. Тело клетки (перикарион) содержит ядро. От перикариона отходят отростки. Один из них — аксон, все другие — дендриты. Справа и сверху вниз: функциональные зоны нервной клетки — рецептивная (дендритная), аксон (область проведения ПД), концевых разветвлений аксона (пресинаптическая). Дендриты — ветвящиеся отростки, заканчивающиеся вблизи от тела нейрона. Дендриты имеют многочисленные шипики, значительно увеличивающие их поверхность. В функциональном отношении дендриты — рецепторная зона (или постсинаптическая область) регистрации сигналов от других нервных клеток. Поверхностная мембрана дендритов и перикариона одного нейрона головного мозга образует до 20 000 синаптических контактов с другими нейронами. Другими словами, в поверхностную мембрану, покрывающую дендриты и перикарион, встроено множество рецепторов к нейромедиаторам, секретируемым другими нервными клетками. Тело нейрона содержит ядро, комплекс Гольджи, гранулярную эндоплазматическую сеть, митохондрии, лизосомы, элементы цитоскелета. Синтез белка происходит только в перикарионе. Интегративная зона — тело нейрона вместе с дендритами и областью отхождения аксона, в этой зоне интегрируются локальные изменения МП, происходящие в местах синаптических контактов. Аксонный холмик (начальный сегмент аксона) — место, где начинается аксон и происходит генерация ПД. Аксон — длинный отросток, как правило, не ветвящийся. Объём аксона может достигать 99% суммарного объёма нейрона. Длина аксона может быть значительной — десятки сантиметров. Основной функцией аксона является проведение ПД. Аксонный транспорт. Поскольку в аксоне и нервных окончаниях практически нет рибосом, необходимые для деятельности всей нервной клетки белки синтезируются в перикарионе, а затем транспортируются по аксону посредством аксонного транспорта. Терминали аксона (концевые разветвления) принимают участие в образовании синапсов с другими нейронами, мышечными и секреторными клетками. Терминали заканчивается утолщениями, содержащими синаптические пузырьки (везикулы). Классификации нейронов Существует множество классификаций нервных клеток, так как нейроны отличаются по размерам и форме перикариона, числу отростков, их синаптическим связям, характеру ветвления дендритов, электрофизиологическим характеристикам, химии нейромедиаторов, позиции в нейронных сетях и множеству других характеристик. В зависимости от класса выполняемой функции выделяют афферентные (чувствительные, сенсорные), эфферентные (двигательные, моторные) и вставочные нервные клетки (ассоциативные нейроны, или интернейроны). Афферентные нейроны проводят возбуждение от рецепторов периферических органов в структуры ЦНС. Эфферентные нейроны осуществляют передачу сигналов от ЦНС к органам-эффекторам (мышцам и железам). Ассоциативные нейроны проводят возбуждение между нейронами. Морфологическая классификация. По количеству отростков нейроны разделяют на псевдоуниполярные, биполярные и мультиполярные. Псевдоуниполярные нейроны имеют один короткий отросток, который разделяется на некотором расстоянии от сомы на два длинных — дендрит и аксон. К псевдоуниполярным относятся нейроны сенсорных ганглиев спинного мозга. Биполярные нейроны имеют один дендрит и один аксон. Этот вид нейронов встречается в периферическом отделе зрительного, обонятельного и слухового анализаторов. Мультиполярные нейроны имеют один аксон и несколько дендритов, это наиболее распространённый вид нейронов. К ним относятся мотонейроны спинного мозга. Исходя из характера электрогенеза в различных зонах нервных клеток (рецептивная и интегративная зоны, начальный сегмент аксона, аксон, терминали аксона), предложена универсальная классификация (рис. 5–2, см. также рис. 5–1). Эта классификация учитывает направление (вход и выход) сигналов в пределах одной нервной клетки, но находящейся в составе нейронных сетей или цепочек, а также характер мембранного электрогенеза (изменения МП в рецептивной и пресинаптической зонах и передача ПД по аксону). 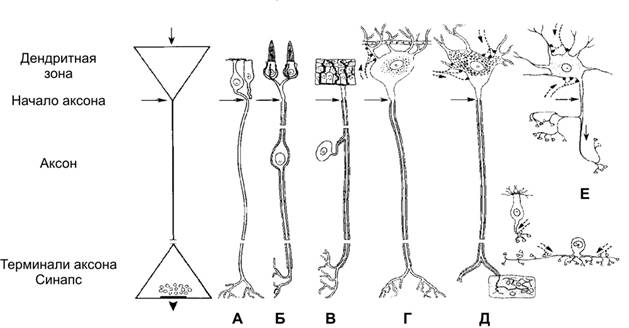 Рис. 5–2. Универсальная классификация частей различных типов нейронов [11]. Дендриты — рецепторная зона — возбуждающий или тормозящий вход. Перикарион с дендритами — интегративная зона. Аксон — проводящая возбуждение (ПД) часть нейрона, его концевые разветвления образуют синапсы (выход) с соседними клетками. А–Г — афферентные (чувствительные) нейроны; Д — мотонейрон; Е — интернейроны. мембранный потенциал покоя Мембранный потенциал (МП) покоя — разность электрических потенциалов между внутренней и наружной поверхностью мембраны в состоянии покоя (при отсутствии раздражителей) — рассмотрен в главе 2 (раздел «Мембранный электрогенез»). МП покоя нервной клетки составляет –70 мВ, и его величина достаточна стабильна. Изменения величины МП (рис. 5–3) описывают специальными терминами: деполяризация — уменьшение величины МП; реполяризация — возвращение величины МП к исходному уровню; гиперполяризация — увеличение МП по сравнению с исходным уровнем. Все изменения МП в дендритной зоне нервной клетки известны как локальные ответы, затухающие на протяжении нескольких микрометров от места возникновения. В этой зоне постоянно возникают десятки и сотни локальных ответов (постсинаптические потенциалы), суммирование которых в виде изменений МП достигает области начала аксона (аксонного холмика). Помимо локальных ответов, возбудимые мембраны нервных клеток в области аксона могут генерировать потенциалы действия (ПД, спайки, нервные импульсы, рис. 5–3 и 5–4). Существенное значение при генерации ПД имеет представление о пороге раздражения. 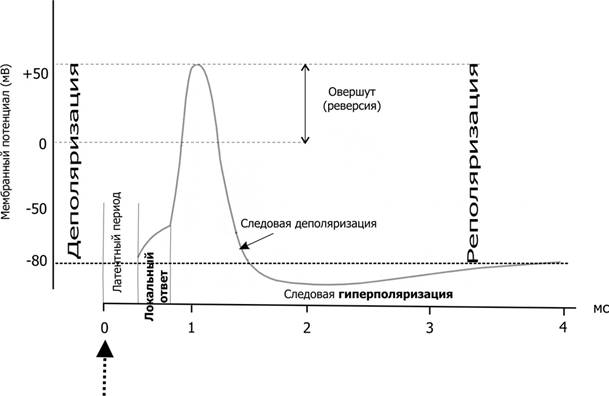 Рис. 5–3. Изменения мембранного потенциала и потенциал действия. Вертикальная стрелка в нижней части рисунка — момент появления раздражающего стимула, на отметке –80 мВ — исходный уровень МП.  Рис. 5–4. Мембранный потенциал и потенциал действия. На отметке –70 мВ — исходный уровень МП; значение порога показано стрелкой в левой части рисунка, приведены записи подпороговых изменений МП. Законы раздражения нервной клетки Генерация ПД подчиняется законам «всё или ничего», «сила–время» и аккомодации. Закон «всё или ничего». При достижении пороговой силы раздражающего стимула дальнейшее увеличение его интенсивности или продолжительности раздражения не изменяет характеристик ПД. Такая форма реакции нервной клетки на раздражение получила название «всё или ничего» (либо ПД возникает, либо нет). Закон силы–времени. Эффект раздражителя на нервную клетку зависит не только от силы раздражителя, но и от времени, в течение которого он действует; так, чем больше сила тока, тем меньше времени он должен действовать, чтобы возник процесс возбуждения, и наоборот. Соотношение силы и длительности действия раздражителя может быть выражено в виде гиперболической кривой (рис. 5–5). 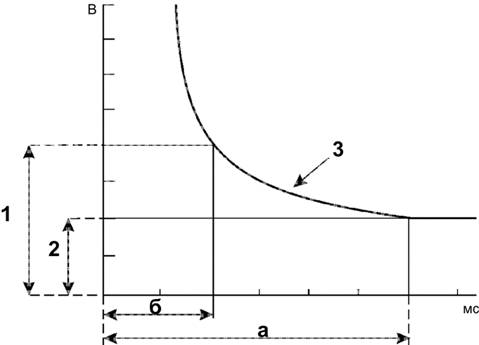 Рис. 5–5. Кривая «силы–времени» [21]. 1 — реобаза (минимальная сила раздражителя, способная вызвать ПД); 2 — удвоенная реобаза; 3 — кривая «силы-времени»; а — полезное время (минимальное время, в течение которого ток, равный одной реобазе, вызывает генерацию ПД); б — хронаксия (минимальное время, в течение которого ток, равный двум реобазам, вызывает ПД). Из графика на рисунке 5–4 видно, что усиление раздражителя приводит к уменьшению минимального времени раздражения, но не бесконечно. При очень малых значениях длительности стимула кривая «силы-времени» становится почти параллельной к оси ординат. Точно также невозможно вызвать генерацию ПД длительным, но очень малым по амплитуде раздражающим стимулом. Закон аккомодации (рис. 5–6). Раздражающее действие тока зависит не только от амплитуды электрического сигнала, но и от скорости (крутизны) его нарастания во времени: чем быстрее это нарастание, тем сильнее выражено раздражающее действие тока. При действии медленно нарастающей силы раздражителя генерации ПД не происходит, так как возбудимая клетка «приспосабливается» к действию этого раздражителя, что и получило название аккомодации. 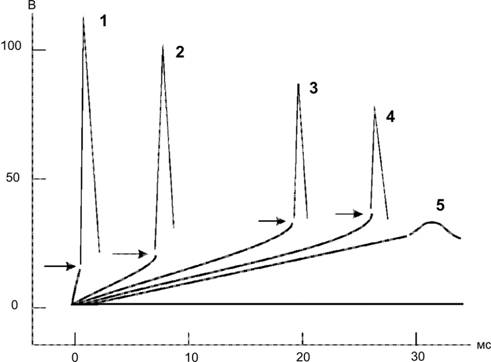 Рис. 5–6. Аккомодация мембраны нервной клетки к действию токов различной крутизны нарастания [21]. На графике показано изменение критического уровня деполяризации и амплитуды ПД (1-5) при раздражении нервной клетки линейно нарастающими токами различной крутизны. Изменения возбудимости во время потенциала действия. В ходе развития ПД происходят изменения возбудимости мембраны нервной клетки. Абсолютно рефрактерный период. Во время фазы деполяризации и большей части фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течение которого даже сверхпороговое раздражение не способно вызвать ПД. Этот феномен связан с инактивацией большинства Na+ каналов. Относительно рефрактерный период. В конце фазы реполяризации, а также во время следовой гиперполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители. Это связано со значительным реполяризующим действием выходящих калиевых токов. Наличие рефрактерности ограничивает частоту генерации ПД. Физиологическое значение рефрактерности заключается в создании условий для своевременного и полного осуществления восстановительных процессов в нервной клетке. Феномен рефрактерности лежит в основе понятия о функциональной подвижности, или лабильности (Н.Е. Введенский). Лабильность — максимально возможная частота генерации ПД для данного типа возбудимой клетки. Лабильность большинства нейронов составляет приблизительно 400 ПД/с, а у интернейронов спинного мозга доходит до 1000 ПД/с. В нормальных условиях рефрактерность предохраняет нервные клетки от излишне частой генерации ПД. Мутации, гипоксия, механические травмы и другие патологические воздействия приводят к значительным изменениям возбудимости нейронов. Такие нейроны или группы нейронов являются потенциальными источниками возникновения пароксизмальных состояний ЦНС, эпилептических припадков и других неврологических расстройств. Проведение возбуждения по нервным волокнам Нервные волокна — аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских клеток в периферических нервах. Нервные волокна подразделяют на 2 типа — безмиелиновые и миелиновые. Основная функция нервных волокон — проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна (рис. 5–8) и существенно зависит от диаметра нервных волокон. 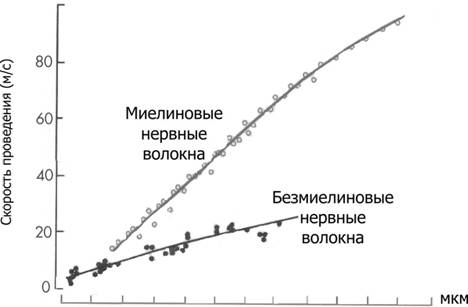 Рис. 5–8. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра [4]. Скорость проведения пропорциональна диаметру нервного волокна и в миелиновых волокнах выше, чем в безмиелиновых. Безмиелиновые нервные волокна (рис. 5–9А). В покое мембрана аксона (осевого цилиндра) поляризована — положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется, и наружная поверхность мембраны приобретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны. Такое проведение известно как электротоническое, а проведение ПД — своего рода «эстафета», в которой каждый участок мембраны является сначала раздражаемым, а затем раздражающим. ПД возникает за счёт увеличения проводимости через потенциалозависимые Na+ каналы, встроенные в аксолемму с плотностью около 110–120 каналов на 1 мкм2. Появление так называемых рефрактерных каналов (рефрактерное состояние мембраны после прохождения ПД) предупреждает распространение возбуждения в обратном направлении. Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5–2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения ПД (см. рис. 5–8). 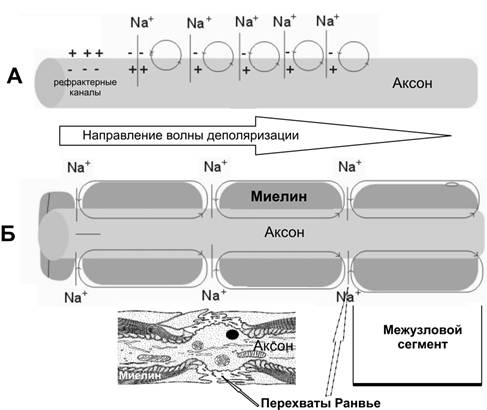 Рис. 5–9. Проведение возбуждения в нервных волокнах [7]. А — безмиелиновое волокно (электротоническое проведение), Б — миелиновое волокно (скачкообразное проведение). Миелин, полностью окружая аксон в межузловых промежутках, выступает в роли электрического изолятора, а межклеточная жидкость в перехватах Ранвье — проводник. Миелиновое нервное волокно (рис. 5–9Б) состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания собственной плазматической мембраны. Миелин прерывается через регулярные промежутки (от 0,2 до 2 мм) концентрической щелью шириной около 1 мкм, это узлы, или перехваты Ранвье. Таким образом, межузловые сегменты аксона, расположенные между соседними перехватами Ранвье, содержат миелин — электрический изолятор, не позволяющий проходить через него локальным токам, поэтому ПД возникают только в перехватах Ранвье. Другими словами, ПД перемещается вдоль нервного волокна скачками, от одного перехвата Ранвье к другому перехвату (скачкообразное проведение). Плотность потенциалозависимых Na+ каналов аксолеммы в перехватах Ранвье — до 2000 на 1 мкм2 (в перикарионе — 50–70, в начальном сегменте аксона — 2000, в межузловых сегментах Na+ каналы практически отсутствуют). В силу высокой плотности Na+ каналов перехваты Ранвье характеризуются высокой возбудимостью, а локальные токи достаточно велики для возбуждения соседнего перехвата. Локальные токи текут от перехвата к перехвату (через внеклеточную жидкость кнаружи от миелина и через аксоплазму внутри аксона) с минимальными потерями. Скорость проведения ПД в миелиновых волокнах в десятки раз выше, чем в наиболее «быстрых» безмиелиновых аксонах. Энергозатраты нервного волокна на проведение ПД относительно невелики, поскольку возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% общей поверхности мембраны аксона. Поэтому даже после длительных ритмических пачек ПД трансмембранный градиент концентраций ионов практически не изменяется. В физиологических условиях ПД движутся в одном направлении от места раздражения (ортодромное проведение). ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения. Проведение в противоположном направлении (антидромное проведение) возможно при травматическом поражении нервных волокон и в редких случаях (аксон–рефлекс). Нарушение миелинизации нервных волокон приводит к нарушениям проводимости (демиелинизирующие заболевания). При разрушении миелиновой оболочки происходит резкое снижение скорости и надёжности проведения возбуждения по нервам. Наиболее распространённым среди демиелинизирующих заболеваний является множественный склероз, проявляющийся различными параличами и потерей чувствительности. Законы проведения возбуждения Бездекрементное проведение возбуждения. Амплитуда ПД в различных участках нерва одинакова, то есть проведение возбуждения по нервному волокну осуществляется без затухания (бездекрементно). Таким образом, кодирование информации осуществляется не за счёт изменения амплитуды ПД, а путём изменения их частоты и распределения во времени. Изолированное проведение возбуждения. Нервные стволы обычно образованы большим количеством нервных волокон, однако ПД, идущие по каждому из них, не передаются на соседние. Эта особенность нервных волокон обусловлена: наличием оболочек, окружающих отдельные нервные волокна и их пучки (в результате образуется барьер, предупреждающий переход возбуждения с волокна на волокно); сопротивлением межклеточной жидкости (жидкость, находящаяся между волокнами, имеет гораздо меньшее сопротивление току, чем мембрана аксонов; поэтому ток шунтируется по межволоконным пространствам и не доходит до соседних волокон). Физиологическая и анатомическая целостность. Необходимым условием проведения возбуждения является не только его анатомическая целостность, но и нормальное функционирование мембраны нервного волокна (физиологическая целостность). В клинике широко применяют различные ЛС, нарушающие физиологическую целостность нервных волокон. Так, эффекты местных анестетиков (новокаин, лидокаин, и др.) основаны на блокаде потенциалозависимых Na+ каналов. Нарушение физиологической целостности чувствительных нервных волокон вызывает анестезию (потерю чувствительности). |