Нейрофизиология Задания. Нейрофизиология ПЛз-19. Тема 1 биоэлектрические явления
 Скачать 0.65 Mb. Скачать 0.65 Mb.
|

|
ПЛз-19 37.03.01 Психология Дисциплина: Нейрофизиология Дата: Тема 1: БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ Цель: изучить способность живых клеток переходить из состояния физиологического покоя в состояние активности. Потенциал покоя Между наружной поверхностью клетки и ее протоплазмой в состоянии покоя существует разность потенциалов порядка 60—90 мв, причем поверхность клетки заряжена электроположительно по отношению к протоплазме. Эту разность потенциалов принято называть мембранным потенциалом покоя, или просто потенциалом покоя. Точное его измерение возможно только с помощью микроэлектродов, предназначенных, для внутриклеточного отведения. Микроэлектрод представляет собой микропипетку, т. е. тонкий капилляр, вытянутый из стеклянной трубки. Диаметр его кончика около 0,5 мк. Микропипетку заполняют солевым раствором (3 М КСl), погружают в него металлический электрод и соединяют с электроизмерительным прибором — осциллографом с усилителем постоянного тока. Микроэлектрод устанавливают над исследуемым объектом, например мышцей, а затем с помощью микроманипулятора — прибора, снабженного микрометрическим винтом, вводят внутрь клетки. Второй электрод обычных размеров прикладывают к поверхности мышцы или же погружают его в раствор Рингера, в котором находится исследуемый объект. 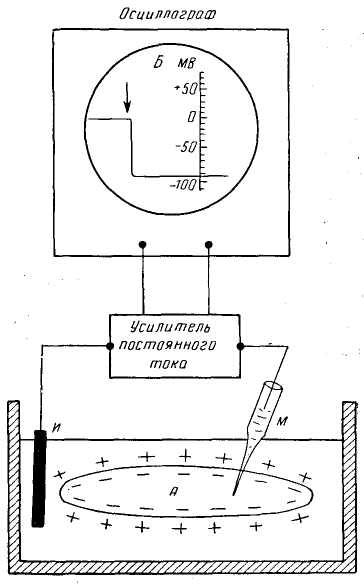 Рис 8. Измерение мембранного потенциала мышечного волокна (А) с помощью внутриклеточного микроэлектрода (схема). М — микроэлектрод; И — индифферентный электрод. Луч на экране осциллографа (Б) показывает, что до прокола микроэлектродом мембраны разность потенциалов между М и И была равна нулю. В момент прокола (показан стрелкой) обнаружена разность потенциалов, указывающая, что внутренняя сторона мембраны заряжена электроотрицательно по отношению к ее наружной поверхности. Как только микроэлектрод прокалывает покрывающую клетку мембрану, луч осциллографа сразу отклоняется вниз от своего исходного положения и устанавливается на новом уровне (рис. 8), обнаруживая тем самым существование скачка потенциала между поверхностью и содержимым клетки. Наличие разности потенциалов между наружной поверхностью клетки и ее содержимым может быть обнаружено и без помощи микроэлектродов. Для этого достаточно нанести поперечный разрез на нерв или мышцу и приложить отводящие электроды таким образом, чтобы один из них касался места разреза, а второй — неповрежденной поверхности. В этом случае электроизмерительный прибор покажет, что между указанными участками ткани протекает ток (ток покоя), причем неповрежденный участок оказывается заряженным электроположительно по отношению к месту разреза. Однако такой способ отведения не позволяет измерять полную разность потенциалов между наружной поверхностью и внутренним содержимым клетки, так как жидкость, омывающая ткань с поверхности и находящаяся в межклеточных щелях, шунтирует регистрирующую систему. Поэтому измеряемая разность потенциалов между поврежденным и неповрежденным участками ткани не превышает обычно 30—50 мв. Происхождение потенциала покоя В 1902 г. Бернштейном была развита мембранно-ионная теория, которая модифицирована и экспериментально обоснована Ходжкиным, Хаксли и Катцем (1949—1952) и в настоящее время пользуется широким признанием. Согласно этой теории, биоэлектрические потенциалы обусловлены неодинаковой концентрацией ионов К+, Na+, Cl- внутри и вне клетки и различной проницаемостью для них поверхностной мембраны. Протоплазма нервных и мышечных клеток содержит в 30—50 раз больше ионов калия, в 8—10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Препятствием для быстрого выравнивания этой разности концентраций является тончайшая (около 100 А) плазматическая мембрана, покрывающая живые клетки. Представления о молекулярной структуре этой мембраны строятся на основании данных, полученных методами электронной микроскопии, диффракции рентгеновых лучей, химического анализа, а также при изучении электрических характеристик этой мембраны. Клеточные мембраны обладают избирательной проницаемостью: одни вещества пропускают, другие не пропускают. В частности, мембрана легкопроницаема для жирорастворимых веществ, проникающих через липидный слой; большинство мембран пропускает воду. Анионы органических кислот не проходят через мембрану, но имеются каналы, избирательно пропускающие ионы К+, Na+, Ca2+, Сl. При действии нервных импульсов проницаемость мембраны нейрона для различных ионов изменяется, это обеспечивает движение ионов согласно концентрационному и электрическому градиентам, что выражается в возникновении возбуждающих и тормозных потенциалов. Проницаемость мембраны для различных катионов также неодинакова и закономерно изменяется при разных функциональных состояниях ткани. В покое мембрана нервных волокон примерно в 25 раз более проницаема для ионов К+, чем для ионов Na+, а при возбуждении натриевая проницаемость примерно в 20 раз превышает калиевую проницаемость мембраны. Для того чтобы понять механизм возникновения мембранного потенциала покоя, рассмотрим модельный опыт. Правую половину сосуда (рис. 9), разделенного искусственной полупроницаемой мембраной, поры которой свободно пропускают положительно заряженные ионы К+ и не пропускают отрицательно заряженных ионов SO4-, наполняют концентрированным раствором K2SO4, а левую половину наполняют также раствором K2SO4, но меньшей концентрации. Вследствие существования концентрационного градиента ионы К+ начнут диффундировать через мембрану преимущественно из правой половины сосуда (где их концентрация равна C1) в левую (с концентрацией С2). Соответственно отрицательно заряженные анионы SO4-, для которых мембрана непроницаема, будут концентрироваться в правой половине сосуда у поверхности мембраны. Своим отрицательным зарядом они электростатически будут удерживать ионы К+ на поверхности мембраны слева. В результате мембрана поляризуется: между двумя ее поверхностями возникает разность потенциалов. Если теперь в правую и левую половины сосуда опустить электроды, электроизмерительный прибор обнаружит наличие разности потенциалов; при этом раствор с меньшей концентрацией ионов K2SO4, в который происходит преимущественно диффузия положительно заряженных ионов К+, приобретает положительный заряд по отношению к раствору с большей концентрацией K2SO4. 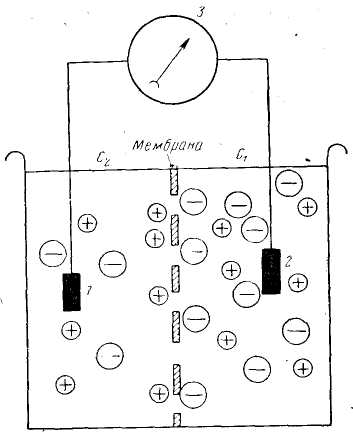 Рис. 9. Возникновение разности потенциалов на искусственной мембране, разделяющей растворы K2SO4 разной концентрации (C1и С2). Мембрана избирательно проницаема для катионов К+ (маленькие кружки) и не пропускает анионы SO4-(большие кружки). 1, 2— электроды, опущенные в раствор; 3 — электроизмерительный прибор. Роль обмена веществ в поддержании потенциала покоя («натрий-калиевый насос») Несмотря на то что скорость диффузии ионов Na+ и К+ через мембрану в покое мала, разность их концентрации вне клетки и внутри нее должна была бы в конечном итоге полностью выровняться, если бы в клетке не существовало специального механизма, который обеспечивает активное выведение («выкачивание») из протоплазмы проникающих в нее ионов Na+ и введение («нагнетание») ионов К+. Этот механизм получил образное название натрий-калиевого насоса. Для того чтобы сохранялась ионная асимметрия, натрий-калиевый насос должен против градиента концентраций выкачивать из клетки ионы Na+ и нагнетать в нее ионы К+ и, следовательно, совершать определенную работу. Непосредственным источником энергии для работы насоса является расщепление богатых энергией, «макроэргических», фосфорных соединений — аденозинтрифосфата (АТФ), которое происходит под влиянием фермента — аденозинтрифосфатазы, локализованной в мембране и активируемой ионами Na+ и К+. Торможение активности этого фермента, вызываемое некоторыми веществами, например оуабаином, приводит к нарушению работы насоса. К такому же результату приводит и угнетение процессов синтеза АТФ в клетке. Так, при отравлении нервных волокон монойодацетатом, выключающим процессы гликолиза, при воздействии цианидов, тормозящих дыхание, вместе с прекращением синтеза АТФ прекращается и работа натрий-калиевого насоса. В результате протоплазма отравленного волокна обогащается натрием и теряет калий. Прямым следствием такого выравнивания ионных концентраций по обе стороны мембраны является снижение или даже полное исчезновение потенциала покоя. Если же с помощью микроканюли в отравленное гигантское волокно ввести некоторое количество АТФ, активный транспорт ионов Na+ и К+ восстанавливается. Из этого можно сделать вывод, что обмен веществ является источником энергии, необходимой для поддержания в живой клетке того неравенства ионных концентраций, от которого зависит потенциал покоя. Механизм возникновения потенциала действия Если действие раздражителя на клеточную мембрану приводит к возникновению потенциала действия (ПД), далее сам процесс развития ПД вызывают фазовые изменения проницаемости клеточной мембраны, что обеспечивает быстрое движение иона Na+ в клетку, а иона К+ - из клетки. Величина мембранного потенциала при этом сначала уменьшается, а затем снова восстанавливается до исходного уровня. На экране осциллографа отмеченные изменения мембранного потенциала представляют в виде пикового потенциала - ПД. Он возникает вследствие накопленных и поддерживаемых ионными насосами градиентов концентраций ионов внутри и вне клетки, т.е. за счет потенциальной энергии в виде электрохимических градиентов разных ионов. Если заблокировать процесс выработки энергии, но после исчезновения градиентов концентраций ионов (устранение потенциальной энергии) клетка генерировать ПД не будет. Рассмотрим фазы ПД: Фаза деполяризации. При действии деполяризующего раздражителя на клетку (медиатор, электрический ток) вначале уменьшение мембранного потенциала (частичная деполяризация) происходит без изменения проницаемости мембраны для ионов. Когда деполяризация достигает примерно 50% пороговой величины (порогового потенциала), возрастает проницаемость ее мембраны для иона Na+, причем в первый момент сравнительно медленно. Естественно, что скорость входа ионов Na+ в клетку при этом невелика. В этот период, как и во время всей фазы деполяризации, движущей силы, обеспечивающей вход иона Na+ в клетку, являются концентрационный и электрический градиенты. Напомним, что клетка внутри заряжена отрицательно (разноименные заряды притягиваются друг к другу), а концентрация ионов Na+ вне клетки в 10-12 раз больше, чем внутри клетки. При возбуждении нейрона повышается проницаемость его мембраны и для ионов Са+, но его ток в клетку значительно меньше, чем ионов Na+. Условием, обеспечивающим вход иона Na+ в клетку и последующий выход иона К+ из клетки, является увеличение проницаемости клеточной мембраны, которая определяется состоянием воротного механизма ионных Na- и К-каналов. Длительность пребывания электроуправляемого канала в открытом состоянии носит вероятностный характер и зависит от величины мембранного потенциала. Суммарный ток ионов в любой момент определяется числом открытых каналов клеточной мембраны. Воротный механизм Na-каналов расположен на внешней стороне клеточной мембраны (Na+ движется внутрь клетки), воротный механизм К-каналов - на внутренней (К+ движется из клетки наружу). Фаза инверсии. После исчезновения ПП вход Na+ в клетку продолжается, поэтому число продолжительных ионов в клетке превосходит число отрицательных, заряд внутри клетки становится положительным, снаружи - отрицательным. Процесс перезарядки мембраны представляет собой 2-ю фазу ПД - фазу инверсии. Теперь электрический градиент препятствует входу Na+ внутрь клетки (положительные заряды отталкиваются друг от друга), проводимость Na+ снижается. Тем не менее, некоторый период (доли миллисекунды) ионы Na+ продолжают входить в клетку, об этом свидетельствует продолжающееся нарастание ПД. Это означает, что концентрационный градиент, обеспечивающий движение ионов Na+ в клетку, сильнее электрического, препятствующего входу ионов Na+ в клетку. Во время деполяризации увеличивается проницаемость ее и для ионов Са2+, они также идут в клетку, но в нервных клетках роль ионов Са2+ в развитии ПД мала. Таким образом, вся восходящая часть пика ПД обеспечивается в основном входом ионов Na+ в клетку. Примерно через 0,5-1 мс. после начала деполяризации рост ПД прекращается вследствие закрытия ворот Na-каналов и открытия ворот К-каналов, т.е. увеличения проницаемости для ионов К+. Поскольку ионы К+ находятся преимущественно внутри клетки, они, согласно концентрационному градиенту, быстро выходят из клетки, вследствие чего в клетке уменьшается число положительно заряженных ионов. Заряд клетки начинает возвращаться к исходному уровню. В фазу инверсии выходу ионов К+ из клетки способствует также электрический градиент. Ионы К+ выталкиваются положительным зарядом из клетки и притягиваются отрицательным зарядом снаружи клетки. Так продолжается до полного положительного заряда внутри клетки - до конца фазы инверсии. Амплитуда ПД складывается из величины ПП (мембранный потенциал покоящейся клетки) и величины фазы инверсии - около 20 мв. Если мембранный потенциал в состоянии покоя клетки мал, то амплитуда ПД этой клетки будет небольшой. Фаза реполяризации. В этой фазе проницаемость клеточной мембраны для ионов К+ все еще высока, ионы К+ продолжают быстро выходить из клетки согласно концентрационному градиенту. Клетка снова внутри имеет отрицательный заряд, а снаружи - положительный, поэтому электрический градиент препятствует выходу К+ из клетки, что снижает его проводимость, хотя он продолжает выходить. Это объясняется тем, что действие концентрационного градиента выражено значительно сильнее действия электрического градиента. Таким образом, вся нисходящая часть пика ПД обусловлена выходом иона К+ из клетки. Нередко в конце ПД наблюдается замедление деполяризации, что объясняется уменьшением проницаемости клеточной мембраны для ионов К+ и замедлением выхода их из клетки вследствие закрытия ворот К-каналов. Другая причина замедления тока ионов К+ связана с возрастанием положительного потенциала наружной поверхности клетки и формированием противоположно направленного электрического градиента. Главную роль в возникновении ПД играет ион Na+, входящий в клетку при повышении проницаемости клеточной мембраны и обеспечивающий всю восходящую часть пика ПД. При замене иона Na+ в среде на другой ион, например холин, или в случае блокировки Na-каналов тетродотоксином, ПД в нервной клетке не возникает. Однако проницаемость мембраны для иона К+ предотвратить тетраэтиламмонием, то мембрана после ее деполяризации реполяризуется гораздо медленнее, только за счет медленных неуправляемых каналов (каналы утечки ионов), через которые К+ будет выходить из клетки (Рис.10). Роль ионов Са2+ в возникновении ПД в нервных клетках незначительна, в некоторых нейронах она существенна, например в дендритах клеток Пуркинье мозжечка. Следовые явления в процессе возбуждения клетки. Эти явления выражаются в гиперполяризации или частичной деполяризации клетки после возвращения мембранного потенциала к исходной величине. 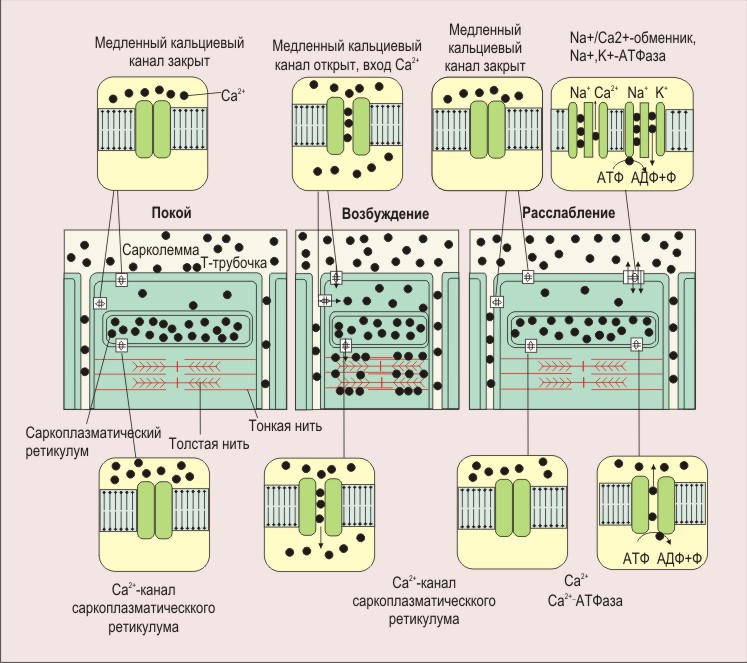 Рис. 10 Состояние К-Nа каналов при возбуждении Классификация ионных каналов. Классифицируют ионные каналы по нескольким признакам. 1. По возможности управления функцией различают неуправляемые (каналы утечки ионов) и управляемые каналы. Через неуправляемые каналы ионы перемещаются постоянно, но медленно, естественно, при наличии электрохимического градиента, как и в случае быстрого перемещения ионов по управляемым каналам. Последние могут быть быстрыми и медленными. Потенциал действия в нейроне возникает в основном вследствие активации быстрых Na- и К-каналов. Управляемые каналы имеют ворота с механизмами их управления, поэтому ионы через них могут проходить только при открытых воротах. 2. В зависимости от стимула, активирующего или инактивирующего управляемые ионные каналы, основными каналами нейронов ЦНС являются потенциал-чувствительные и хемо-чувствительные каналы. При взаимодействии медиатора (лиганда) с рецепторами хемо-чувствительного канала, расположенного на поверхности клеточной мембраны, может происходить открытие его ворот, поэтому хемо-чувствительный канал называют также рецепторуправляемым каналом. Лиганд - это биологически активное вещество или фармакологический препарат, активирующий или блокирующий рецептор. Открытие хемо-чувствительных каналов происходит в результате конформационных изменений рецепторного комплекса. Ворота потенциалзависимых каналов открываются и закрываются при изменении величины мембранного потенциала. Поэтому в конструкции их воротного механизма должны быть частицы, несущие электрический заряд. 3. В зависимости от селективности различают ионоселективные каналы, пропускающие только один ион, и каналы, не обладающие селективностью. В нейронах имеются Na-, K-, Са- и С1-селективные каналы. Есть каналы, пропускающие несколько ионов, например Na+, K+ и Са2+, т.е. не обладающие селективностью. Наиболее высока степень селективности потенциал-чувствительных (потенциалзависимых) каналов, несколько ниже она у хемо-чувствительных (рецепторзависимых) каналов, постсинаптических мембран, через каналы которых могут одновременно проходить ионы Na+и К+. 4. Для одного и того же иона может быть несколько видов каналов. Наиболее важными из них для формирования биопотенциалов являются следующие. Каналы для ионов К+. Калиевые неуправляемые каналы покоя (каналы утечки), через которые постоянно выходит К+ из клетки, что является главным фактором в формировании мембранного потенциала (потенциала покоя). Потенциал-чувствительные управляемые К-каналы, сравнительно медленно активирующиеся при возбуждении клетки в фазу деполяризации с последующим ускорением активации, что обеспечивает быстрый выход ионов К+ из клетки и реполяризацию ее. Каналы для ионов Na+. Они бывают быстрые и медленные (утечки). Быстрые Na-каналы потенциал-чувствительны, быстро активируются при уменьшении мембранного потенциала, что обеспечивает вход иона Na+ в клетку во время ее возбуждения (восходящая часть). Затем эти каналы быстро инактивируются. Медленные неуправляемые Na-каналы - каналы утечки, через которые ион Na+ постоянно диффундирует в клетку и переносит с собой другие молекулы, например глюкозу, аминокислоты, молекулы-переносчики. Таким образом, Na-каналы утечки обеспечивают вторичный транспорт веществ и участие ионов Na+ в формировании мембранного потенциала. Ионные каналы блокируются специфическими веществами и фармакологическими препаратами. Новокаин, например, как местный анестетик снимает болевые ощущения потому, что он, блокируя Na-каналы, прекращает проведение возбуждения по нервным волокнам. Основные свойства нервной клетки 1. Возбудимость - это свойство нейрона генерировать потенциал действия на раздражение. Исходя из этого определения, необходимо отметить следующее. Возбудимость является частным случаем наиболее общего свойства всех клеток - раздражимости. К возбудимым клеткам относятся только те, которые генерируют потенциал действия. Это мышечные и нервные клетки. Нередко к возбудимым тканям относят и «железистую ткань». Однако для этого нет оснований, поскольку железистой ткани нет, имеются различные железы и железистый эпителий как вид ткани. В процессе активной деятельности железы в ней действительно регистрируются биоэлектрические явления, так как железа как орган состоит из различных клеток: соединительной ткани, эпителиальной, мышечной. ПД проводится по мембранам нервных и мышечных клеток, с его помощью передается информация и обеспечивается управление деятельностью клеток организма. Невозбудимыми тканями являются эпителиальная и соединительная (собственно соединительная, ретикулярная, жировая, хрящевая, костная и гематопоэтические ткани в совокупности с кровью). Клетки этих тканей не генерируют потенциалы действия при действии на них раздражителя. Раздражитель - это любое изменение внешней или внутренней среды организма, воспринимаемое клетками и вызывающее ответную реакцию. В зависимости от природы раздражители делят на физические (электрические, механические, температурные, световые) и химические, в зависимости от степени чувствительности клеток к тому или иному раздражителю - на адекватные и неадекватные. Адекватный раздражитель - это такой раздражитель, к которому клетка обладает наибольшей чувствительностью вследствие наличия специальных структур, воспринимающих этот раздражитель. Адекватным раздражителем нейронов являются медиаторы и электрические импульсы. Из всех видов раздражителей в эксперименте часто используют электрический раздражитель, поскольку он является универсальным, его легко дозировать по силе, длительности, частоте и крутизне нарастания силы стимула. 2. Проводимость - это способность ткани и клетки проводить возбуждение. Процессы возбуждения и торможения нервных клеток (электрические явления) обеспечивают выполнение их функций. Следует отметить, что все клетки и ткани организма обладают раздражимостью - это общее свойство живой материи активно изменять характер своей жизнедеятельности при действии раздражителя. Реакции отдельных клеток, тканей на действие раздражителя могут быть весьма разнообразными: изменение интенсивности обмена веществ, рН, проницаемости клеточной мембраны, движение ионов, электрические явления, ускорение клеточного деления и роста, структурно-функциональные реакции (например, выделение секрета железистыми клетками или сокращение мышечных клеток). Степень выраженности возможных изменений может быть различной, она зависит от вида клеток или ткани, которые подвергаются действию раздражителя. Таким образом, возбудимость - это частный вид раздражимости. Задание 1. Обозначьте изменения возбудимости мембраны при развитии потенциала действия на указанном ниже графике 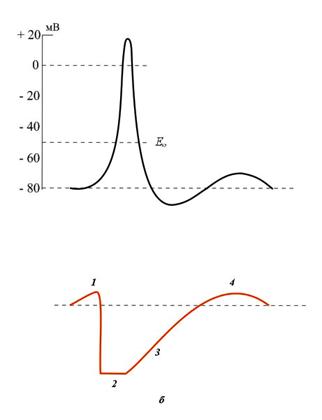 Задание 2.Опишите и объясните явления и механизмы, представленные на графиках Закон «Все или ничего» 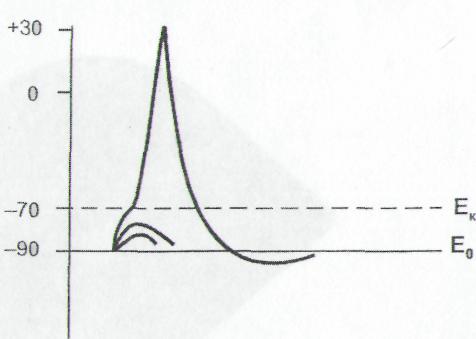 ________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________ Закон:«Сила—длительность» 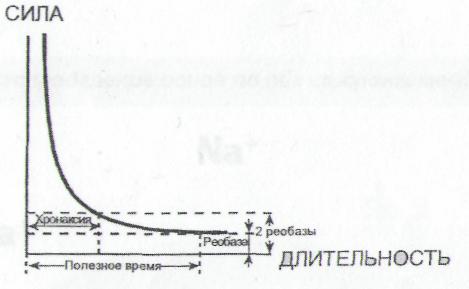 __________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________ Закон Градиента раздражения (Аккомодация) _  ______________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________ ______________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________Задание3. Используя знания о способности живых клеток переходить из состояния физиологического покоя в состояние активности, заполните таблицу: Таблица 4.
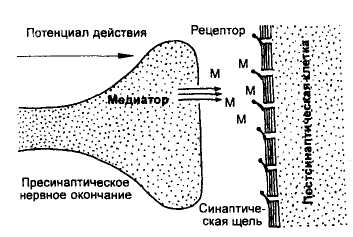
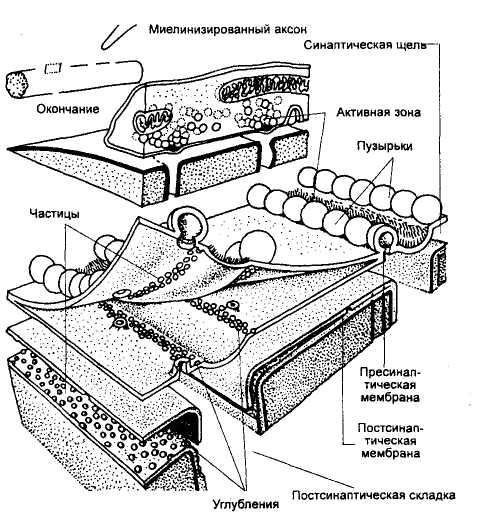
Тема 2: Физиология синапсов Цель: изучить ультраструктуру нервно-мышечного синапса и особенности передачи химического сигнала в синапсе. Синапсами называют специализированные контакты между нервными клетками или между нервными и эффекторными клетками, используемые для передачи сигналов. Синапсы можно классифицировать: по их местоположению и принадлежности соответствующим клеткам — нервно-мышечные, нейро-нейрональные, а среди последних — аксосоматические, аксодендритические синапсы; по знаку их действия — возбуждающие и тормозящие; по способу передачи сигналов — электрические (в которых сигналы передаются электрическим током) и химические, в которых передатчиком, трансмиттером сигнала, или посредником, медиатором, является то или иное физиологически активное вещество. Существуют и смешанные — электрохимические — синапсы. Во всех синапсах содержатся такие компоненты, как пресинаптическая мембрана, постсинаптическая мембрана и разделяющая их синаптическая щель. Химические синапсы возбуждающего действия. В отличие от электрических химические синапсы (рис.11) имеют относительно широкую синаптическую щель, составляющую 20-50 нм, и высокое сопротивление синаптических мембран. Поперечных каналов, связывающих клетки, здесь нет. Другим характерным признаком химического синапса является наличие в пресинаптической нервной терминали большого числа пузырьков — пресинаптических везикул диаметром около 50 нм. Эти везикулы заполнены медиатором — химическим передатчиком (раздражителем).
Экзоцитозные пузырьки образуются в аппарате Гольджи. В пузырьки упаковываются белки, образовавшиеся в рибосомах эндоплазматического ретикулума. Низкомолекулярные вещества (медиаторы, некоторые гормоны) попадают в везикулы преимущественно с помощью вторичного транспорта. Пузырьки транспортируются посредством сократительного аппарата клетки, состоящего из нитей актина и миозина и микротрубочек, к клеточной мембране, сливаются с ней, а содержимое клетки выделяется во внеклеточную среду. Энергия АТФ расходуется на деятельность сократительного аппарата клетки. Процесс слияния везикул с клеточной мембраной активируется фосфолипидом, лизолецитином и уровнем внутриклеточных ионов Са2+. Например, поступление ионов Са2+ в нервное окончание обеспечивает выделение медиатора через пресинаптическую мембрану в синаптическую щель. Классическим представителем группы химических синапсов является возбуждающий нервно-мышечный синапс скелетной мускулатуры позвоночных (рис.12), действующий с помощью медиатора ацетилхолина (Ах). В нем, как и в любом химическом синапсе, пресинаптический ПД не может петлей своего тока возбудить постсинаптическую клетку; Кпздесь не превышает 0,001. Ток, который выходит через пресинаптическую мембрану, здесь почти целиком уходит через широкую синаптическую щель мимо постсинаптической клетки, обладающей значительным входным сопротивлением. Небольшая же часть этого тока, которая все же входит в постсинаптическую клетку, вызывает лишь ничтожное изменение ее МП.
Суть работы химического синапса состоит в следующем (рис.12, 13). Пресинаптический ПД работает как инициатор нейросекреторного акта. При развитии ПД терминали (а также и при искусственной деполяризации) в нее из среды входят ионы Са2+. Это стимулирует практически синхронный выброс медиатора в синаптическую щель из 100-200 пресинаптических везикул, каждая из которых содержит порцию — квант Ах. Большинство исследователей полагают, что этот выброс медиатора осуществляется путем экзоцитоза — опорожнения везикулы в синаптическую щель. Существует и другая точка зрения: квант медиатора аккумулирован в особых участках пресинаптической мембраны — операторах, которые и выбрасывают Ах в щель, а везикулы — это лишь депо Ах и других веществ. Выход медиатора сильно зависит от величины деполяризации терминали. Эта крутая зависимость объясняет резкое снижение выхода медиатора при падении амплитуды ПД терминали. В нормальных условиях в ответ на нервный импульс высвобождается около миллиона молекул Ах (в каждом кванте — везикуле — их примерно 104). Медиатор диффундирует к постсинаптической мембране, где для него существуют рецепторы (холинорецепторы — Хр). При взаимодействии Ах и Хр в последних открываются проницаемые для Na+ и К+ ионные каналы с d ≈ 0,65 нм. Так как холинорецепторов и, соответственно, каналов много, сопротивление постсинаптической мембраны сильно падает, что приводит к ее частичной деполяризации, т. е. к развитию возбуждающего постсинаптического потенциала (ВПСП). 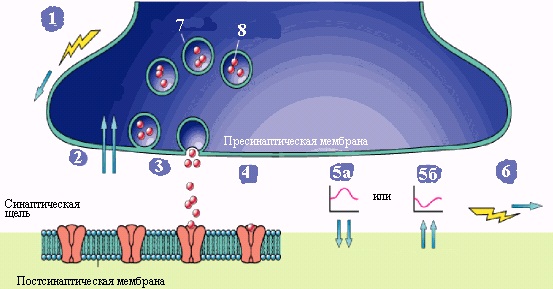 Рис. 13. Схема передачи химического сигнала в синапсе. Задание 1. Опишите образования и процессы, обозначенные цифрами на рисунке 13 (смотри пример под цифрой 1): 1. Потенциал действия (ПД) приходит по аксону в пресинаптическую область и деполяризует пресинаптическую мембрану. 2. _________________________________________________________________________________________________________________________________________________________ 3. __________________________________________________________________________________________________________________________________________________________ 4. __________________________________________________________________________________________________________________________________________________________ 5. _________________________________________________________________________________________________________________________________________________________ 6. __________________________________________________________________________________________________________________________________________________________ 7. __________________________________________________________________________________________________________________________________________________________ 8. ________________________________________________________________________________________________________________________________________________________ Запомните еще два важных свойства синапсов: Первое: синапсы выполняют функцию усилителей электрических (нервных) сигналов при передаче ПД с постсинаптической мембраны на постсинаптическую. В чем заключается суть усиления: один ПД инициирует выход из пресинаптической мембраны 100 везикул с 6000-8000 молекул нейротрансмиттера внутри каждой везикулы. Такой мощный химический сигнал воздействует на сравнительно небольшую по площади часть постсинаптической мембраны, на которой расположены рецепторы белковой природы (протеины), связывающиеся с молекулами нейротрансмиттера. Они вновь, но с усилением трансформируют химический сигнал в электрический на постсинаптической мембране. Усиление в химическом синапсе осуществляется также за счет участия вторичных посредников- активаторов метаболических процессов в клетках. Описанные теоретические знания широко применяются в практической медицине. Например, действие многих современных лекарственных средств основано на управлении химическими сигналами в синапсах: усилении (облегчении условий передачи) или подавлении (вплоть до полной блокады). Второе: синапсы (постсинаптическая мембрана) могут изменять чувствительность к действию нейротрансмиттеров: малое количество медиатора при длительном действии на постсинаптическую мембрану повышает ее чувствительность - явление сенситизации; высокая концентрация медиатора при длительном действии на постсинаптическую мембрану снижает ее чувствительность - явление десенситизации. Эта функция синапсов называется синаптической пластичностью и составляет основу таких процессов, как память и обучение. Общая характеристика синаптических медиаторов |