Биология 1-2 вариант .Филогенез. 1-3 вариант Биология. Вариант 1 1 Профаза i профаза первого деления очень сложная и состоит из 5 стадий 2N4c лептотена
 Скачать 125.95 Kb. Скачать 125.95 Kb.
|

|
Вариант 1 1) Профаза I — профаза первого деления очень сложная и состоит из 5 стадий:2N4C Лептотена, или лептонема — упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются). Зиготена, или зигонема — происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация. Пахитена, или пахинема — (самая длительная стадия) — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер — обмен участками между гомологичными хромосомами. Диплотена, или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток. Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой. К концу профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления, разрушаются ядерная мембрана и ядрышки. Генетический материал — 2n4c (n — число хромосом, c — число молекул ДНК). Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки. Генетический материал — 2N4C. Анафаза I — микротрубочки сокращаются, биваленты делятся, и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе. У каждого полюса генетический материал n2c, во всей клетке 2N4C Телофаза I — хромосомы деспирализуются и появляется ядерная оболочка.1N2C 2) Митохондрия. Функции митохондрий как органелл аэробных эукариотических клеток – синтез молекул АТФ (аденозинтрифосфата) из АДФ. Поскольку АТФ является универсальным источником энергии для всех процессов в клетке, идущих с потреблением энергии, то говорят, что митохондрии выполняют функцию энергообеспечения, или энергообразования. 1.Наружная мембрана 2.Внутренняя мембрана 3.Кристы 4.Грибовидные тела(?) 5.Матрикс 6.Кольцевая ДНК 3) 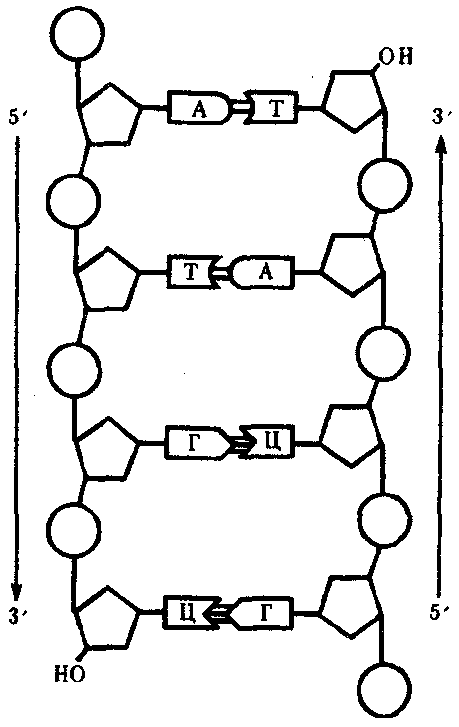 Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы (фосфодиэфирные связи). Антипараллельность цепей ДНК: противоположная направленность двух нитей двойной спирали ДНК; одна нить имеет направление от 5' к 3', другая - от 3' к 5'. Каждая цепь ДНК имеет определенную ориентацию Комплемента́рность (в химии, молекулярной биологии и генетике) — взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими (комплементарными) фрагментами молекул или их структурных фрагментов вследствие супрамолекулярных взаимодействий (образование водородных связей, гидрофобных взаимодействий, электростатических взаимодействий заряженных функциональных групп и т. п.). В случае нуклеиновых кислот — как олиго- так и полинуклеотидов, азотистые основания нуклеотидов способны вследствие образования водородных связей формировать парные комплексы аденин—тимин (или урацил в РНК) и гуанин—цитозин при взаимодействии цепей нуклеиновых кислот. Такое взаимодействие играет ключевую роль в ряде фундаментальных процессов хранения и передачи генетической информации: репликации ДНК, обеспечивающей передачу генетической информации при делении клетки, транскрипции ДНК в РНК при синтезе белков, кодируемых ДНК гена, хранении генетической информации в двухцепочечной ДНК и процессах репарации ДНК при её повреждении. 4) 1) фрагмент двухцепочечной молекулы ДНК: -ЦАЦТАТЦЦАГАТАГА- -ГТГАТАГГТЦТАТЦТ-; 2) последовательность иРНК: - ЦАЦУАУЦЦАГАУАГА-; 3) последовательность аминокислот: -гис-тирпро-асп-арг 5)Под трансляцией в биологии понимают синтез из аминокислот полипептидов, который протекает в цитоплазме на рибосомах при участии 1) мРНК в качестве матрицы, 2) тРНК в качестве переносчика аминокислот, а также 3) ряда белковых факторов, выполняющих каталитическую функцию на разных этапах процесса.Активация аминокислотАктивация аминокислот необходима, так как только в таком состоянии они способны соединяться с тРНК и позже образовывать между собой пептидные связи. В цитоплазме клеток всегда находятся свободные (не соединенные с другими веществами) аминокислоты. Специфичные ферменты в присутствии АТФ преобразуют аминокислоту в аминоациладенилат, который уже способен соединяться с тРНК. Существует класс ферментов – аминоацил-тРНК-синтетазы, – которые активируют аминокислоты, используя при этом энергию АТФ. Каждая аминокислота активируется своим ферментом, после чего присоединяется только к своей тРНК. Образуется комплекс аминокислоты с тРНК – аминоацил-тРНК (аа-тРНК). Инициация трансляции Инициация трансляции включает следующие последовательно протекающие при участии факторов инициации этапы: Присоединение 5'-конца мРНК к малой субъединице рибосомы. При этом стартовый кодон (AUG) размещается в недостроенном (из-за отсутствия большой субъединицы) P-сайте рибосомы. Комплекс аа-тРНК с соответствующим антикодоном присоединяется к стартовому кодону мРНК. У эукариот кодон AUG кодирует аминокислоту метионин, у прокариот — формил-метионин. Позже эти стартовые аминокислоты вырезаются из готового полипептида. Происходит объединение субъединиц рибосом, в результате чего достраиваются их P- и A-сайты. 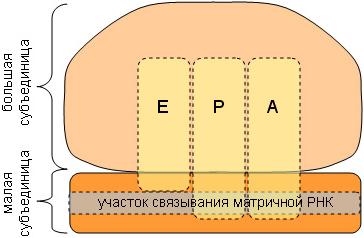 Схема строения рибосомы (A, P, E — участки-сайты для молекул тРНК) Схема строения рибосомы (A, P, E — участки-сайты для молекул тРНК)Таким образом, на этапе инициации происходит распознавание рибосомой стартового кодона и подготовка к началу синтеза. Образующаяся связь между рибосомой и мРНК обратима, мРНК после синтеза полипептида может быть отсоединена от рибосомы. В последствии мРНК используется еще раз или разрушается специальными ферментами. Стартовый кодон AUG отличается от других таких же кодонов в середине мРНК тем, что перед ним находится кэп и определенные нуклеотидные последовательности. Именно благодаря им AUG распознается как стартовый. (Это касается в основном эукариот.) Элонгация трансляции На этом этапе происходит непосредственный синтез полипептидной цепочки. Процесс элонгации состоит из множества циклов. Один цикл элонгации — это присоединение одной аминокислоты к растущей полипептидной цепочке. Уже на этапе инициации P-сайт рибосомы оказывается занятым первой тРНК, несущей аминокислоту метионин. В первом цикле элонгации в A-сайт рибосомы заходит второй комплекс aa-тРНК. Это будет та тРНК, чей антикодон комплементарен следующему (за стартовым AUG) кодону. A(аминоацил)- и P(пептидил)-сайты располагают комплексы аа-тРНК так, что между аминокислотами протекает химическая реакция, и образуется пептидная связь. После этого первая (находящаяся в P-сайте) тРНК освобождается от своей аминокислоты. В результате последняя оказывается связанной только со второй аминокислотой пептидной связью. Вторая аминокислота связана со второй тРНК, находящейся в A-сайте. Рибосома перемещается по нити мРНК на один триплет. При этом первая т-РНК оказывается в E-сайте (exit) рибосомы, после чего покидает ее. Вторая т-РНК, связанная с двумя аминокислотами, переходит в P-сайт. A-сайт освобождается для поступления третьего комплекса аа-тРНК. Следующие циклы элонгации протекают аналогично первому. Когда A-сайт освобождается, в него может зайти аа-тРНК, чей антикодон комплементарен кодону мРНК, находящемся в этот момент в A-сайте. Терминация трансляции Терминация — это завершения синтеза полипептидной цепочки и ее отделение. Терминация наступает, когда рибосома встречает один из терминирующих кодонов (UAA, UAG, UGA), для которых не существует своих тРНК. Эти участки мРНК распознаются специальными белками — факторами терминации. ВАРИАНТ 2 1) Стадии, или фазы, второго мейотического деления: Профаза II. Разрушение ядерных оболочек, формирование веретена деления.1N2C Метафаза II. Хромосомы располагаются в экваториальной плоскости, к ним присоединяются нити веретена деления. Причем таким образом, что к каждой центромере присоединяются две нити — одна с одного полюса, другая — с другого.1N2C Анафаза II. Хроматиды каждой хромосомы разделяются в области центромер, и каждая из пары сестринских хроматид уходит к своему полюсу.2N2C Телофаза II. Формирование ядер, раскручивание хромосом, деление цитоплазмы.1N1C 2) Комплекс Гольджи. Имеет одну мембрану Функции аппарата Гольджи: 1) накопление белков, липидов, углеводов, 2) модификация поступивших органических веществ, 3) «упаковка» в мембранные пузырьки белков, липидов, углеводов, 4) секреция белков, липидов, углеводов, 5) синтез углеводов и липидов, 6) место образования лизосом. 3) Ген — это участок ДНК, несущий какую-либо целостную информацию — о строении одной молекулы белка или одной молекулы РНК Свойства гена: 1) Дискретность действия, так как он определяет возможность развития отдельного качества, присущего данной клетке или организму. 2) Специфичное действие, ввиду того, что в гене заключается информация об аминокислотной последовательности определенного полипептида. 3) Плейотропное действие. В некоторых случаях одна и та же нуклеотидная последовательность может детерминировать синтез не одного, а нескольких полипептидов. Это наблюдается в случае альтернативного сплайсинга у эукариот и при перекрывают генов у фагов и прокариот (Определяя возможность транскрибирования мРНК для синтеза конкретной полипептидной цепи, ген характеризуется дозированностью действия, т.е. количественной зависимостью результата его экспрессии от дозы соответствующего аллеля этого гена) 4) 5) Транскри́пция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.Транскрипция катализируется ферментом ДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5'- к 3'- концу, то есть по матричной цепи ДНК РНК-полимераза движется в направлении 3'->5'Транскрипция состоит из стадий инициации, элонгации и терминации. Инициация транскрипции происходит в специфических участках ДНК – промоторах(Промотор (promoter): последовательность ДНК , которая связывает РНК-полимеразу и служит отправной точкой транскрипции;) , - и сама состоит из нескольких стадий. В отличие от ДНК-полимераз , РНКП способна самостоятельно осуществлять инициацию и начинать синтез РНК в отсутствие затравки.Инициация требует наличия субстратов РНК-полимеразы - нуклеозидтрифосфатов - и заключается в образовании первых нескольких звеньев цепи РНК.При синтезе первых 9 рибонуклеотидов этот фрагмент выбрасывается и затем транскрипция начинается вновь и идет до конца.Энхансер — небольшой участок ДНК, способный связываться с факторами транскрипции, при этом увеличивая уровень транскрипции гена или группы генов.Сайленсер (англ. Silencer) — последовательность ДНК, с которой связываются белки-репрессоры (факторы транскрипции). Связывание белков-репрессоров с сайленсерами приводит к понижению или к полному подавлению синтеза РНК ферментом ДНК-зависимой РНК-полимеразой. Элонгация-Переход от инициации к элонгации сопровождается разрывом связей между ферментом, промотором, факторами инициации транскрипции.Момент перехода РНК-полимеразы от инициации транскрипции к элонгации точно не определен.Происходит наращивание цепи, в результате которой происходит расплетение ДНК, транскрибция восст. Струк. ДНК с выталкиванием РНК. ИЛИ Факторы элонгации повышают активность РНК-полимеразы и облегчают расхождение цепей ДНК.Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК. Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади - восстановление двойной спирали ДНК. Терминация-Раскручивание двойной спирали ДНК в области сайта терминации делает его доступным для фактора терминации. Завершается синтез РНК в строго определенных участках матрицы - терминаторах (сайты терминации).Сигнал об окончании транскрипции закодирован в мРНК и наз. Терминатор.он обр. в самой РНК.есть два механизма терминации транскрипции: ро-зависимый механизм, при котором белок Rho (ро) дестабилизирует водородные связи между матрицей ДНК и мРНК, высвобождая молекулу РНК. ро-независимый, при котором транскрипция останавливается, когда только что синтезированная молекула РНК формирует стебель-петлю, за которой расположено несколько урацилов (…УУУУ), что приводит к отсоединению молекулы РНК от матрицы ДНК. Терминация транскрипции у эукариот менее изучена. Она завершается разрезанием РНК, после чего к её 3' концу фермент добавляет несколько аденинов (…АААА), от числа которых зависит стабильность данного транскрипта Вариант 3 1) Профаза I — профаза первого деления очень сложная и состоит из 5 стадий:2N4C Лептотена, или лептонема — упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются). Зиготена, или зигонема — происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация. Пахитена, или пахинема — (самая длительная стадия) — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер — обмен участками между гомологичными хромосомами. Диплотена, или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток. Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой. К концу профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления, разрушаются ядерная мембрана и ядрышки. Генетический материал — 2n4c (n — число хромосом, c — число молекул ДНК). 2) Микротрубочки Микротрубочки — это структуры, в которых 13 протофиламентов, состоящих из гетеродимеров α- и β-тубулина, уложены по окружности полого цилиндра. Внешний диаметр цилиндра около 25 нм, внутренний — около 15 Микротрубочки в клетке используются в качестве «рельсов» для транспортировки частиц. По их поверхности могут перемещаться мембранные пузырьки и митохондрии. Транспортировку по микротрубочкам осуществляют белки, называемые моторными. Это высокомолекулярные соединения, состоящие из двух тяжёлых (массой около 300 кДа) и нескольких лёгких цепей. В тяжёлых цепях выделяют головной и хвостовой домены. Два головных домена связываются с микротрубочками и являются собственно двигателями, а хвостовые — связываются с органеллами и другими внутриклеточными образованиями, подлежащими транспортировке. 3)  5) Информационная РНК - доставляет генетическую информацию от ДНК к рибосоме (месту белкового синтеза). Транспортная РНК - осуществляет перенос аминокислот к рибосомам (месту синтеза белка). Рибосомная РНК - участвует в формировании активного центра рибосомы, а также обеспечивает взаимодействие транспортной РНК и рибосомы.  |