Семинар 4. Вопрос Вопрос Потенциал действия. Ионный механизм возникновения потенциала действия
 Скачать 2.09 Mb. Скачать 2.09 Mb.
|

|
Вопрос 1.  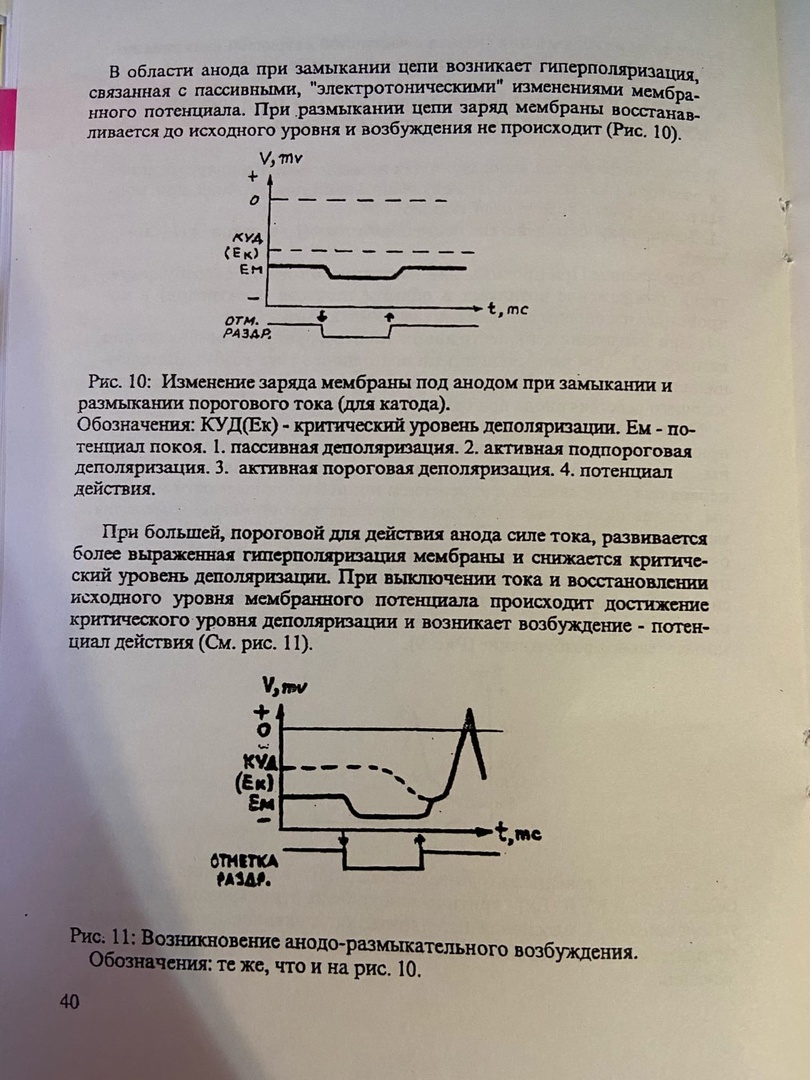    Вопрос 2. Потенциал действия. Ионный механизм возникновения потенциала действия Потенциалом действия (ПД) называют быстрое колебание мембранного потенциала, возникающее при возбуждении нервных, мышечных и некоторых других клеток. В его основе лежат изменения ионной проницаемости мембраны. Амплитуда ПД мало зависит от силы вызывающего его раздражителя, важно лишь, чтобы эта сила была не меньше некоторой критической величины, которая называется порогом раздражения. Возникнув в месте раздражения, ПД распространяется вдоль нервного или мышечного волокна, не изменяя своей амплитуды. В естественных условиях ПД генерируются в нервных волокнах при раздражении рецепторов или возбуждении нервных клеток. Распространение ПД по нервным волокнам обеспечивает передачу информации в нервной системе. Достигнув нервных окончаний, ПД вызывают секрецию химических веществ (медиаторов), обеспечивающих передачу сигнала на мышечные или нервные клетки. В мышечных клетках ПД инициируют цепь процессов, вызывающих сократительный акт. Ионы, проникающие в цитоплазму во время генерации ПД, оказывают регулирующее влияние на метаболизм клетки и, в частности, на процессы синтеза белков, составляющих ионные каналы и ионные насосы. В ПД выделяют следующие фазы (рис. 3): деполяризации; реполяризации. 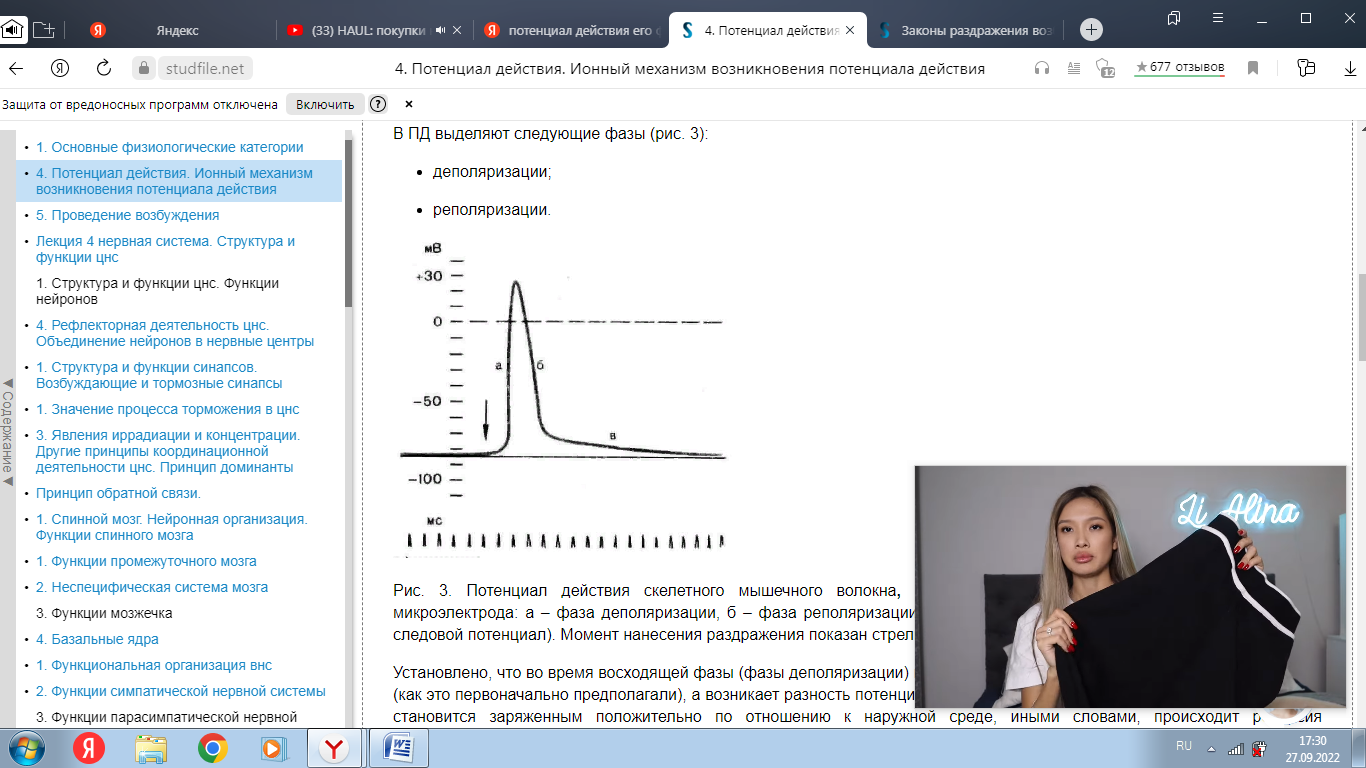 Рис. 3. Потенциал действия скелетного мышечного волокна, зарегистрированный с помощью внутриклеточного микроэлектрода: а – фаза деполяризации, б – фаза реполяризации, в – фаза следовой деполяризации (отрицательный следовой потенциал). Момент нанесения раздражения показан стрелкой. Установлено, что во время восходящей фазы (фазы деполяризации) происходит не просто исчезновение потенциала покоя (как это первоначально предполагали), а возникает разность потенциалов обратного знака: внутреннее содержимое клетки становится заряженным положительно по отношению к наружной среде, иными словами, происходит реверсия мембранного потенциала. Во время нисходящей фазы (фазы реполяризации) мембранный потенциал возвращается к своему исходному значению. Если рассмотреть пример записи ПД в скелетном мышечном волокне лягушки (см. рис. 3), то видно, что в момент достижения пика мембранный потенциал составляет +30 – +40 мВ. Длительность пика ПД у различных нервных и мышечных волокон варьирует от 0,5 до 3 мс, причем фаза реполяризации продолжительнее фазы деполяризации. Изменения мембранного потенциала, следующие за пиком потенциала действия, называют следовыми потенциалами. Различают два вида следовых потенциалов – следовую деполяризацию и следовую гиперполяризацию. Ионный механизм возникновения ПД. Как отмечалось, в состоянии покоя проницаемость мембраны для калия превышает ее проницаемость для натрия. Вследствие этого поток К+ из цитоплазмы во внешний раствор превышает противоположно направленный поток Na+. Поэтому наружная сторона мембраны в покое имеет положительный потенциал по отношению к внутренней. При действии на клетку раздражителя проницаемость мембраны для Na+ резко повышается и становится примерно в 20 раз больше проницаемости для K+. Поэтому поток Na+ из внешнего раствора в цитоплазму начинает превышать направленный наружу калиевый ток. Это приводит к изменению знака (реверсии) мембранного потенциала: внутренняя сторона мембраны в месте возбуждения становится заряженной положительно по отношению к ее наружной поверхности. Указанное изменение мембранного потенциала соответствует восходящей фазе ПД (фазе деполяризации). Повышение проницаемости мембраны для Na+ продолжается лишь очень короткое время. Вслед за этим проницаемость мембраны для Na+ вновь понижается, а для K+ возрастает. Процесс, ведущий к понижению ранее увеличенной натриевой проницаемости мембраны, назван натриевой инактивацией. В результате инактивации поток Na+ внутрь цитоплазмы резко ослабляется. Увеличение же калиевой проницаемости вызывает усиление потока K+ из цитоплазмы во внешний раствор. В итоге этих двух процессов и происходит реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к наружной стороне мембраны. Этому изменению потенциала соответствует нисходящая фаза ПД (фаза реполяризации). Опыты на гигантских нервных волокнах кальмара позволили получить подтверждение правильности натриевой теории возникновения ПД. ПД возникает при деполяризации поверхностной мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению ионов Na внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные изменения на мембране (локальный ответ). При увеличении силы раздражения, когда достигнут порог возбудимости, изменения мембранного потенциала достигают критического уровня деполяризации (КУД). Например, величина потенциала покоя равна -70 мВ, КУД = -50 мВ. Чтобы вызвать возбуждение, надо деполяризовать мембрану до -50 мВ, т.е. на -20 мВ снизить ее исходный потенциал покоя. Только при достижении КУД наблюдается резкое изменение мембранного потенциала, которое регистрируется в виде ПД. Таким образом, основное условие возникновения потенциала действия – это снижение мембранного потенциала до критического уровня деполяризации. В основе рассмотренных изменений ионной проницаемости мембраны при генерации ПД лежат процессы открывания и закрывания специализированных ионных каналов в мембране, обладающих двумя важнейшими свойствами: избирательностью (селективностью) по отношению к определенным ионам; электровозбудимостью, т.е. способностью открываться и закрываться в ответ на изменения мембранного потенциала. Так же как ионные насосы, ионные каналы образованы макромолекулами белков, пронизывающими липидный бислой мембраны. Активный и пассивный ионный транспорт. В процессе восстановления после ПД работа калий-натриевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, благодаря чему нарушенное при возбуждении неравенство концентраций Na+ и K+ по обе стороны мембраны восстанавливается. На работу этого механизма тратится около 70 % необходимой клетке энергии. Таким образом, в живой клетке существует две системы движения ионов через мембрану. Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии (пассивный ионный транспорт). Он ответствен за возникновение потенциала покоя и ПД и ведет в конечном итоге к выравниванию концентрации ионов по обе стороны клеточной мембраны. Второй осуществляется против концентрационного градиента. Он состоит в «выкачивании» ионов натрия из цитоплазмы и «нагнетании» ионов калия внутрь клетки. Этот тип ионного транспорта возможен лишь при условии затраты энергии обмена веществ. Его называют активным ионным транспортом. Он ответствен за поддержание постоянства разности концентраций ионов между цитоплазмой и омывающей клетку жидкостью. Активный транспорт - результат работы натриевого насоса, благодаря которому восстанавливается исходная разность ионных концентраций, нарушающаяся при каждой вспышке возбуждения. Вопрос 3. Законы и меанизмы проведения возбуждения по нервам и нервным волокнам Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью. Проведение возбуждения по нервным волокнам осуществляется по определенным законам. Закон двустороннего проведения возбуждения по нервному волокну.Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу. Закон анатомической ифизиологической целостности нервного волокна.Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности). Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается. Закон изолированногопроведения возбуждения по нервномуволокну.В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервноговолокна на другое, то нормальное функционирование органов было бы невозможно. Возбуждение (потенциал действия) распространяется по нервному волокну без затухания. Периферический нерв практически неутомляем. Механизм проведения возбуждения по нерву.Возбуждение (потенциал действия - ПД) распространяется в аксонах, телах нервных клеток, а также иногда в дендритах без снижения амплитуды и без снижения скорости (бездекрементно). Механизм распространения возбуждения у различных нервных волокон неодинаков. При распространении возбуждения по безмиелиновому нервному волокну механизм проведения включает два компонента: раздражающее действие катэлектротона, порождаемое локальным ПД, на соседний участок электровозбудимой мембраны и возникновение ПД в этом участке мембраны. Локальная деполяризация мембраны нарушает электрическую стабильность мембраны, различная величина поляризации мембраны в смежных ее участках порождает электродвижущую силу и местный электрический ток, силовые линии которого замыкаются через ионные каналы. Активация ионного канала повышает натриевую проводимость, после электротонического достижения критического уровня деполяризации (КУД) в новом участке мембраны генерируется ПД. В свою очередь этот потенциал действия вызывает местные токи, а они в новом участке мембраны генерируют потенциал действия. На всем протяжении нервного волокна происходит процесс новой генерации потенциала действия мембраны волокна. Данный тип передачи возбуждения называется непрерывным. Скорость распространения возбуждения пропорциональна толщине волокна и обратно пропорциональна сопротивлению среды. Проведение возбуждения зависит от соотношения амплитуды ПД и величины порогового потенциала. Этот показатель называется гарантийный фактор(ГФ) и равен 5 - 7, т.е. ПД должен быть выше порогового потенциала в 5- 7 раз. Если ГФ = 1 проведение ненадёжно, если ГФ < 1 проведения нет. Протяженность возбуждённого участка нерва L является произведение времени (длительности) ПД и скорости распространения ПД. Например, в гигантском аксоне кальмара L= 1 мс25 мм/мс = 25 мм. Наличие у миелиновых волоконоболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Вмиелинизированном волокне токи проводятся только в зонах, не покрытых миелином, - перехватах Ранвье, в этих участках и генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением. Распространение ПД в миэлинизированных нервных волокнах осуществляетсясальтаторно- скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Скорость такого способа проведения возбуждения значительно выше, и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов, благодаря чему уменьшается нагрузка на ионный насос. 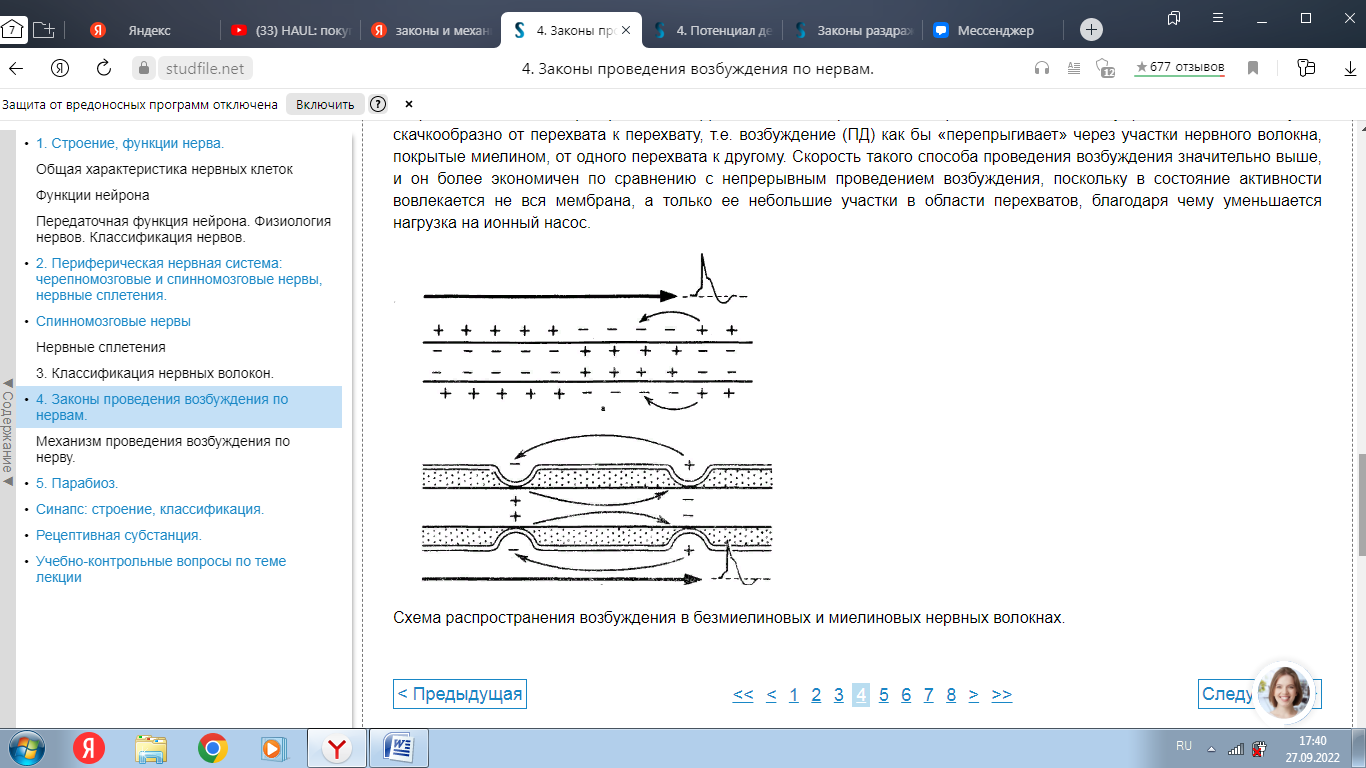 Схема распространения возбуждения в безмиелиновых и миелиновых нервных волокнах. |