Клеточный цикл. 1. Клеточный цикл
 Скачать 1.42 Mb. Скачать 1.42 Mb.
|

|
1.Клеточный цикл представляет собой сложную последовательность событий, предшествующих делению или гибели клетки. В эукариотических клетках этот процесс включает в себя четыре отдельные фазы: фаза Gap 1 (G1); фаза синтеза (S); фаза Gap 2 (G2); митоз (M). Фазы G1, S и G2 клеточного цикла вместе называются интерфазой. Делящаяся клетка проводит большую часть своего времени именно в интерфазе, поскольку при подготовке к делению она растет. Фаза митоза связана с разделением ядерных хромосом с последующим цитокинезом (разделение цитоплазмы на две отдельные клетки). В конце митотического цикла образуются две разные дочерние клетки. Каждая клетка содержит идентичный генетический материал. Время, необходимое для завершения деления клетки зависит от ее типа. К примеру, клетки крови в костном мозге, клетки кожи, клетки желудка и кишечника, делятся быстро и постоянно. Другие клетки делятся при необходимости, заменяя поврежденные или мертвые клетки. К таким типам клеток относятся клетки почек, печени и легких. Другие типы клеток, в том числе нервные клетки, прекращают деление после созревания. Периоды и фазы клеточного цикла Два основные периода клеточного цикла эукариот включат интерфазу и митоз: Интерфаза Во время этого периода клетка удваивает свою цитоплазму и синтезирует ДНК. По оценкам, делящаяся клетка тратит около 90–95% своего времени на интерфазу, которая состоит из следующих 3-х фаз: Фаза G1: промежуток времени до синтеза ДНК. В этой фазе клетка увеличивает свои размеры и количество органелл, подготавливаясь к делению. Клетки животных в этой фазе диплоидны, что означает наличие двух наборов хромосом. S-фаза: этап цикла, в течение которого синтезируется ДНК. В большинстве клеток имеется узкое временное окно, в течение которого происходит синтез ДНК. Содержание хромосом в этой фазе удваивается. Фаза G2: период после синтеза ДНК, но до начала митоза. Клетка синтезирует дополнительные белки и продолжает увеличиваться в размерах. Фазы митоза Во время митоза и цитокинеза содержимое материнское клетки равномерно распределяется между двумя дочерними клетками. Митоз имеет пять фаз: профаза, прометафаза, метафаза, анафаза и телофаза. Профаза: на этой стадии изменения происходят как в цитоплазме, так и в ядре делящейся клетки. Хроматин конденсируется в дискретные хромосомы. Хромосомы начинают мигрировать к центру клетки. Ядерная оболочка ломается и волокна шпинделя образуются на противоположных полюсах клетки. Прометафаза: фаза митоза в эукариотических соматических клетках после профазы и предшествующая метафазе. В прометафазе ядерная мембрана распадается на многочисленные «мембранные везикулы», а хромосомы внутри образуют белковые структуры, называемые кинетохорами. Метафаза: на этом этапе ядерная мембрана полностью исчезает, формируется веретено деление, а хромосомы располагаются на метафазной пластине (плоскость, которая одинаково удалена от двух полюсов клетки). Анафаза: на этой стадии парные хромосомы (сестринские хроматиды) разделяются и начинают двигаться к противоположным концам (полюсам) клетки. Веретено деления, не связанное с хроматидами, вытягивается и удлиняет клетку. Телофаза: на этом этапе хромосомы достигают новых ядер, а генетическое содержание клетки делится поровну на две части. Цитокинез (деление эукариотической клетки) начинается до конца митоза и заканчивается вскоре после телофазы. Цитокинез Цитокинез – процесс разделение цитоплазмы в эукариотических клетках, которые продуцируют различные дочерние клетки. Цитокинез возникает в конце клеточного цикла после митоза или мейоза. При делении клеток животных цитокинез возникает, когда сократительное кольцо образует расщепленную борозду, которая зажимает клеточную мембрану пополам. В растительных клетках строится клеточная пластинка, которая делит клетку на две части. Как только клетка завершит все фазы клеточного цикла, она возвращается в фазу G1 и весь цикл повторяется снова. Клетки организма также способны находится в состояние покоя, которое называется фазой Gap 0 (G0) в любой момент своего жизненного цикла. Они могут оставаться на этой стадии в течение очень длительного периода времени, пока не поступят сигналы к прохождению через клеточный цикл. Клетки, которые содержат генетические мутации, постоянно помещаются в фазу G0, чтобы препятствовать их реплицированию. Когда клеточный цикл идет не так, как надо, нарушается нормальный рост клеток. Могут развиться раковые клетки, которые получают контроль над своими собственными сигналами роста и продолжают размножаться беспрепятственно. Клеточный цикл и мейоз Не все клетки делятся через процесс митоза. Организмы, которые размножаются половым путем, также подвергаются типу клеточного деления, называемого мейозом. Мейоз возникает в половых клетках и аналогичен процессу митоза. Однако после полного клеточного цикла в мейозе образуются четыре дочерние клетки. Каждая клетка содержит половину числа хромосом исходной (родительской) клетки. Это означает, что половые клетки являются гаплоидными клетками. Когда гаплоидные мужские и женские половые клетки объединяются в процессе, называемом оплодотворением, они образуют одну диплоидную клетку, называемую зиготой. 2. Способы деления эукариотических клеток: митоз, мейоз, амитоз Митотический цикл. Митоз Митоз — основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала. Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл. Ниже приводится краткая характеристика фаз цикла. Интерфаза состоит из трех периодов: пресинтетического, или постмитотического, — G1, синтетического — S, постсинтетического, или премитотического, — G2. Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду. Синтетический период (2n 4c) — репликация ДНК. Постсинтетический период (2n 4c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей. Профаза (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом. Метафаза (2n 4c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом. Анафаза (4n 4c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами). Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки. Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др. Мейоз — это особый способ деления эукариотических клеток, в результате которого происходит переход клеток из диплоидного состояния в гаплоидное. Мейоз состоит из двух последовательных делений, которым предшествует однократная репликация ДНК. Первое мейотическое деление (мейоз 1) называется редукционным, поскольку именно во время этого деления происходит уменьшение числа хромосом вдвое: из одной диплоидной клетки (2n 4c) образуются две гаплоидные (1n 2c). Интерфаза 1 (в начале — 2n 2c, в конце — 2n 4c) — синтез и накопление веществ и энергии, необходимых для осуществления обоих делений, увеличение размеров клетки и числа органоидов, удвоение центриолей, репликация ДНК, которая завершается в профазе 1. Профаза 1 (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом, конъюгация гомологичных хромосом и кроссинговер. Конъюгация — процесс сближения и переплетения гомологичных хромосом. Пару конъюгирующих гомологичных хромосом называют бивалентом. Кроссинговер — процесс обмена гомологичными участками между гомологичными хромосомами. Профаза 1 подразделяется на стадии: лептотена (завершение репликации ДНК), зиготена (конъюгация гомологичных хромосом, образование бивалентов), пахитена (кроссинговер, перекомбинация генов), диплотена (выявление хиазм, 1 блок овогенеза у человека), диакинез (терминализация хиазм). Метафаза 1 (2n 4c) — выстраивание бивалентов в экваториальной плоскости клетки, прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом. Анафаза 1 (2n 4c) — случайное независимое расхождение двухроматидных хромосом к противоположным полюсам клетки (из каждой пары гомологичных хромосом одна хромосома отходит к одному полюсу, другая — к другому), перекомбинация хромосом. Телофаза 1 (1n 2c в каждой клетке) — образование ядерных мембран вокруг групп двухроматидных хромосом, деление цитоплазмы. У многих растений клетка из анафазы 1 сразу же переходит в профазу 2. Второе мейотическое деление (мейоз 2) называется эквационным. Интерфаза 2, или интеркинез (1n 2c), представляет собой короткий перерыв между первым и вторым мейотическими делениями, во время которого не происходит репликация ДНК. Характерна для животных клеток. Профаза 2 (1n 2c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления. Метафаза 2 (1n 2c) — выстраивание двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом; 2 блок овогенеза у человека. Анафаза 2 (2n 2с) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами), перекомбинация хромосом. Телофаза 2 (1n 1c в каждой клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток. Биологическое значение мейоза. Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. Являясь основой комбинативной изменчивости, мейоз обеспечивает генетическое разнообразие гамет. Амитоз — прямое деление интерфазного ядра путем перетяжки без образования хромосом, вне митотического цикла. Описан для стареющих, патологически измененных и обреченных на гибель клеток. После амитоза клетка не способна вернуться в нормальный митотический цикл. Амитоз – наиболее простой процесс, чем митоз или мейоз. Амитоз у эукариотов встречается довольно редко и более свойственен прокариотам. Это более быстрый и экономичный процесс, чем митоз. Наблюдается при стремительном восстановлении тканей. Амитозом делятся стареющие клетки и клетки ткани, которые в дальнейшем не будут делиться митотическим способом. Чаще всего это группа клеток, выполняющая строго определённые функции. Амитоз наблюдается: при увеличении корневого чехлика; в клетках эпителия; при росте лука; в рыхлой соединительной ткани; в хрящевой ткани; в мускулатуре; в клетках зародышевых оболочек; при увеличении тканей водорослей; в клетках эндосперма. Основные особенности амитоза, по сравнению с митозом: не сопровождается перестройкой всей клетки; отсутствует веретено деления; не происходит спирализация хроматина; не выявляются хромосомы; отсутствие репликации (удвоения) ДНК; генетический материал распределяется неравномерно; образовавшаяся клетка не способна к митозу. Амитоз может происходить в опухолевых тканях. При неравномерном распределении генетического материала образуются дефектные эукариотические клетки с нарушенными внутриклеточными процессами. Механизм Амитоз – простой и редкий способ деления клеток, который мало изучен. Известно, что амитоз происходит за счёт простой перетяжки (инвагинации) кариолеммы – ядерной оболочки, что приводит к разделению родительской клетки на две части. Во время деления клетка находится в интерфазе, т.е. в состоянии роста и развития, никак не подготавливаясь к делению. Процесс амитоза описан в таблице.
Не всегда при амитозе происходит цитокинез, т. е. деление тела клетки – цитоплазмы со всем её содержим. В этом случае образуется два и более ядра под одной оболочкой (многоядерная клетка), что может приводить к образованию колоний (дрожжи). Значение Амитоз имеет биологическое значение для быстрого восстановления тканей, размножения одноклеточных эукариотических и прокариотических организмов. Амитоз свойственен дрожжам, размножающимся бесполым путём (почкованием), бактериям, лейкоцитам человека. Бактерии и другие прокариоты не имеют ядра. Поэтому амитоз происходит несколько иначе. Сначала удваивается кольцевая ДНК, прикреплённая к складке цитоплазматической мембраны (мезосоме). Затем между двумя закреплёнными на мезосомах ДНК образуется перетяжка, разделяющая клетку пополам. Такое деление часто называют простым или бинарным. 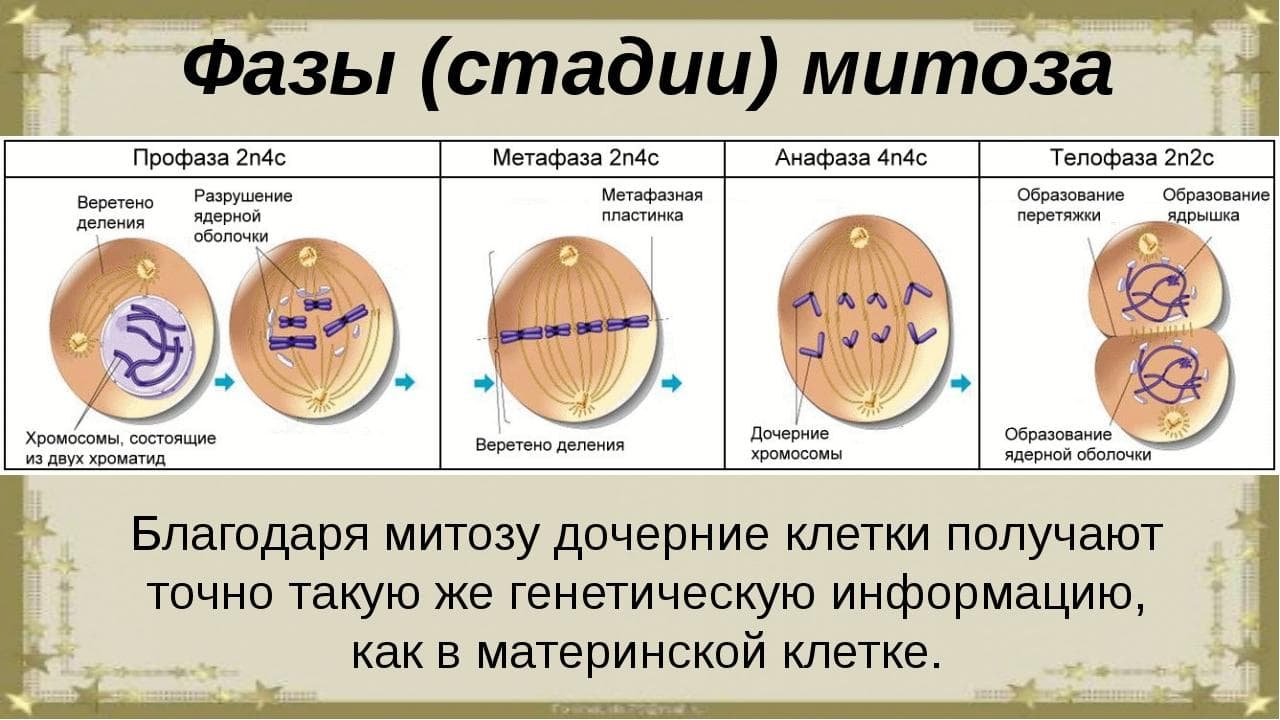 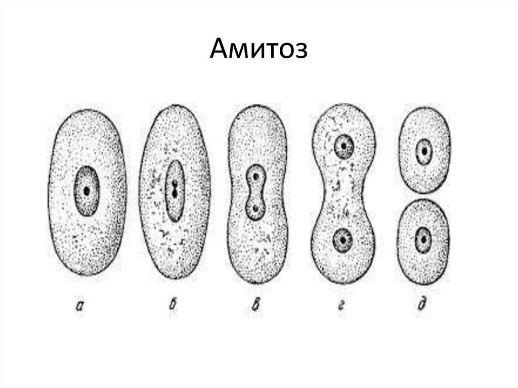 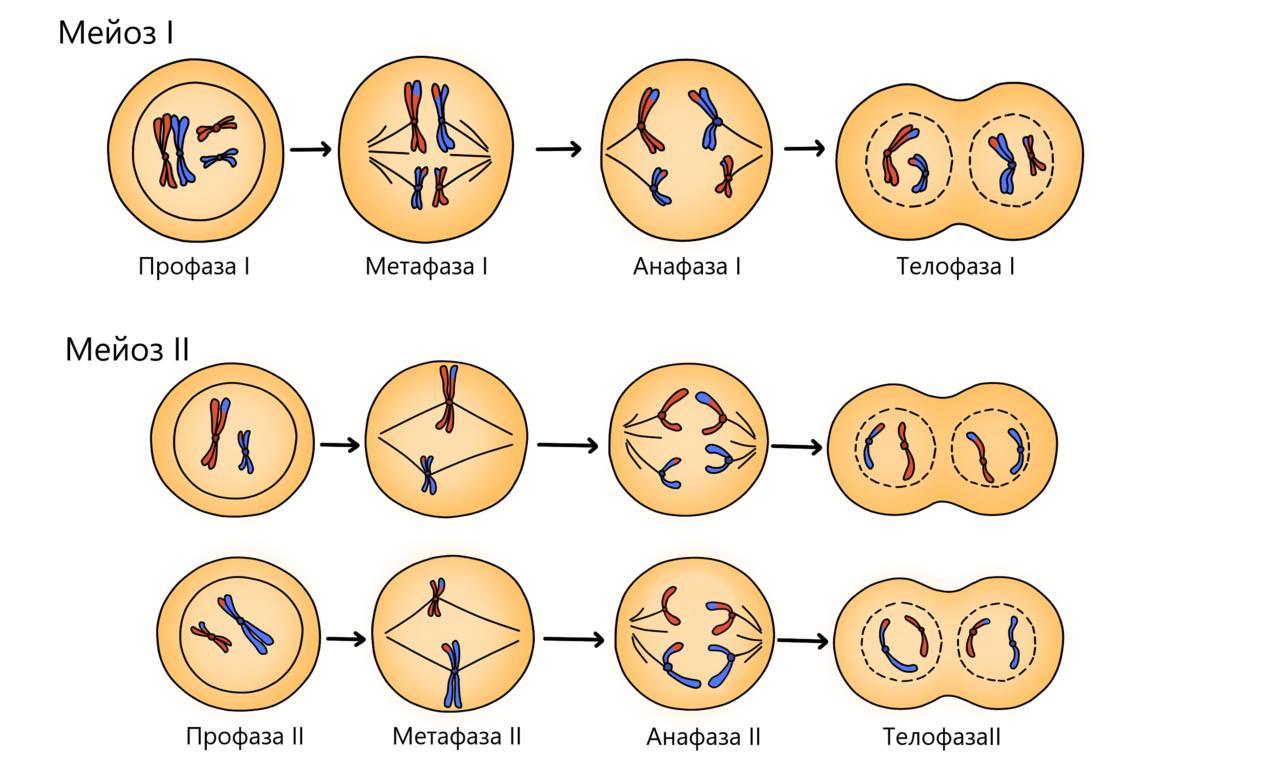 3. Обмен веществ (метаболизм) – совокупность биохимических реакций, протекающих в клетке и обеспечивающих процессы ее жизнедеятельности. В ходе превращения веществ в клетках образуются конечные продукты обмена, которые могут быть токсичными для организма и выводятся из него (например, аммиак). Таким образом, все живые организмы постоянно потребляют из окружающей среды определенные вещества, преобразуют их и выделяют в среду конечные продукты. В зависимости от общей направленности процессов выделяют катаболизм и анаболизм. Анаболизм (ассимиляция) – совокупность химических процессов, направленных на образование и обновление структурных частей клеток, этот процесс имеет второе название – пластический обмен. Фотосинтез: 6Н2О + 6СО2 → С6Н12О6 + 6СО2 ↑ Сюда можно отнести, например, фиксацию азота и биосинтез белка, синтез углеводов из углекислого газа и воды в ходе фотосинтеза, синтез полисахаридов, липидов, нуклеотидов, ДНК, РНК и других веществ. Анаболизм является созидательным этапом обмена веществ, он всегда осуществляется с потреблением энергии и с участием ферментов. Катаболизм (диссимиляция) – совокупность реакций, в которых происходит распад крупных органических молекул до простых соединений с одновременным высвобождением энергии. Катаболизм обеспечивает энергией все процессы, протекающие в клетке, и имеет второе название – энергетический обмен. Дыхание: С6Н12О6 + 6СО2 → 6Н2О + 6СО2 + АТФ 4.Этапы энергетического обмена Рассмотрим энергетический обмен в многоклеточном организме (например, человеческом) и в одноклеточном (например, клетке простейшего). Первый этап – подготовительный. В желудочно-кишечном тракте многоклеточных организмов он осуществляется пищеварительными ферментами. У одноклеточных – ферментами лизосом. На первом этапе происходит расщепление белков до аминокислот, жиров до глицерина и жирных кислот, полисахаридов до моносахаридов, нуклеиновых кислот до нуклеотидов. Этот процесс называется пищеварением. Вся энергия на этом этапе рассеивается в виде тепла. Второй этап – бескислородный (гликолиз). Гликолиз происходит в цитоплазме клеток. Он состоит из нескольких последовательных реакций превращения молекулы глюкозы в две молекулы пировиноградной кислоты (пирувата/ПВК) и две молекулы АТФ: С6Н12O6 + 2АДФ + 2Ф → 2С3Н4O3 + 2АТФ. Остальная энергия рассеивается в виде тепла. В клетках дрожжей и растений (при недостатке кислорода) пируват распадается на этиловый спирт и углекислый газ. Этот процесс называется спиртовым брожением. Энергии, накопленной при гликолизе, слишком мало для организмов, использующих кислород для своего дыхания. Вот почему в мышцах животных, в том числе и у человека, при больших нагрузках и нехватке кислорода образуется молочная кислота (С3Н6O3), которая накапливается в виде лактата. Появляется боль в мышцах. У нетренированных людей это происходит быстрее, чем у людей тренированных Третий этап – кислородный. Состоит из двух последовательных процессов – цикла Кребса, названного по имени Нобелевского лауреата Ганса Кребса, и окислительного фосфорилирования. Его смысл заключается в том, что при кислородном дыхании пируват окисляется до окончательных продуктов – углекислого газа и воды, а энергия, выделяющаяся при окислении, запасается в виде 36 молекул АТФ. (34 молекулы в цикле Кребса и 2 молекулы в ходе окислительного фосфорилирования). Энергия распада органических соединений обеспечивает реакции их синтеза в пластическом обмене. Эволюционно кислородный этап возник после накопления в атмосфере достаточного количества молекулярного кислорода и появления аэробных организмов. Окислительное фосфорилирование, или клеточное дыхание. Происходит на внутренних мембранах митохондрий, в которые встроены молекулы-переносчики электронов и ферментные комплексы АТФ-синтазы. Молекулы-переносчики транспортируют электроны к молекулярному кислороду. Часть энергии рассеивается в виде тепла, а часть расходуется на образование АТФ. Суммарная реакция энергетического обмена: С6Н12O6 + 6O2 → 6СO2 + 6Н2O + 38АТФ. 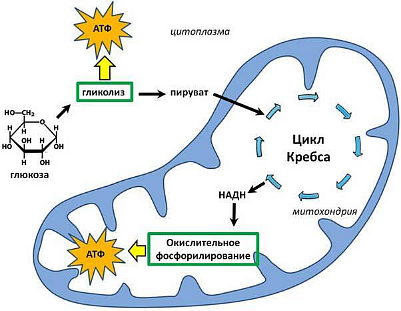 Для удобства запоминания, используйте таблицу с этапами энергетического обмена. 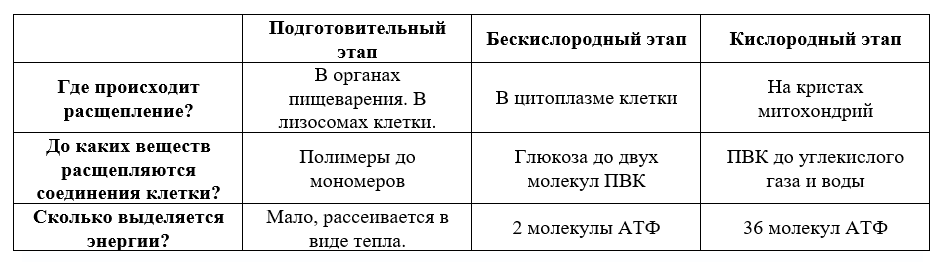 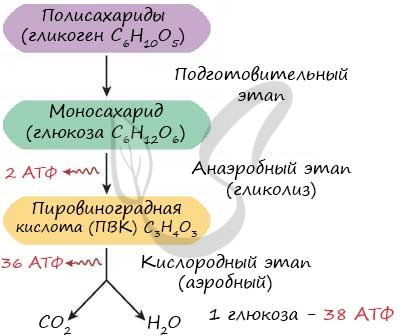 5. Автотрофная ассимиляция Автотрофные организмы способны синтезировать органические вещества из неорганических (СО2 и Н2О). К ним относят зеленые растения и микроорганизмы. В зависимости от того, какой источник энергии используется автотрофными организмами для синтеза органических веществ, их делят на две группы: фототрофы и хемотрофы. Фотосинтез Зеленые растения - фототрофы. Для ассимиляции они используют энергию, выделяемую при окислении неорганических веществ. Зеленые растения имеют в хлоропластах хлорофилл. При участии хлорофилла происходит фотосинтез. Фотосинтез - процесс преобразования солнечной энергии в потенциальную энергию химических связей в органических веществах. Фотосинтез состоит из двух фаз: световой и темновой. 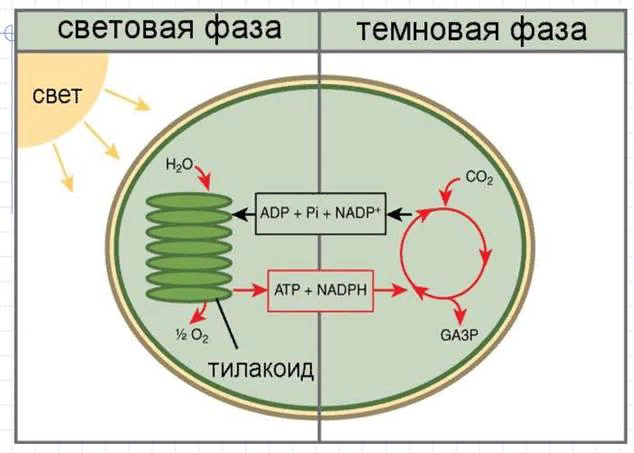 Световая фаза. Под действием света молекула хлорофилла, находящаяся в гранах хлоропласта, получает избыток энергии. Часть этой энергии идет на расщепление (фотолиз) молекулы воды. Ионы водорода присоединяют к себе электрон, превращаются в свободный атом водорода. Водород Н идет на восстановление белка - переносчика НАДФ+ (никотинамидадениндинуклеотид фосфат). НАДФ*Н переходит в строму хлоропласта, где участвует в синтезе углеводов, в частности, глюкозы. Ионы ОН-, отдав электрон, превращаются в свободные радикалы, которые взаимодействуют друг с другом, образуют воду и свободный кислород. Другая часть энергии используется для синтеза АТФ из АДФ. Таким образом, в световую фазу фотосинтеза образуются: 1) богатое энергетическими связями вещество - АТФ; 2) свободный кислород - О2; 3) происходит присоединение Н (водорода) к переносчику, образуется НАДФ*Н. Реакции световой фазы идут без участия ферментов. Темновая фаза. В темновой фазе происходит связывание СО2. В реакциях темновой фазы участвуют молекулы АТФ и атомы водорода, образовавшиеся в процессе фотолиза и связанные с молекулами-переносчиками. Реакции этой фазы происходят в строме хлоропластов при участии ферментов (Цикл Кальвина). Полученные в результате темновой фазы фотосинтеза молекулы моносахарида - глюкозы через ряд ферментативных реакций превращаются в полисахариды. Так энергия солнечного света преобразуется в энергию химических связей сложных органических веществ. 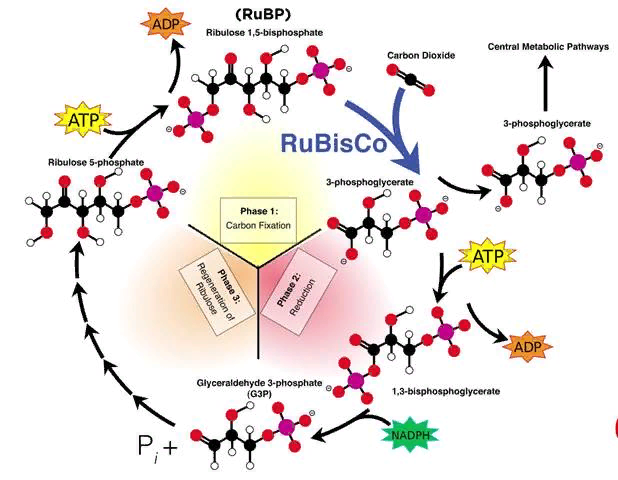 Суммарная реакция фотосинтеза: В результате фотосинтеза образуются органические вещества и кислород атмосферы. Хемосинтез Синтез органических веществ у автотрофных бактерий идет с использованием энергии, выделяющейся при химических реакциях окисления неорганических соединений: сероводорода, серы, аммиака, азотистой кислоты. Источник водорода для восстановления СО2 – вода. Этот процесс называется хемосинтезом. К группе автотрофов-хемосинтетиков относят нитрифицирующие бактерии. Одна группа бактерий получает энергию, необходимую для синтеза органических веществ, в результате реакции окисления аммиака в азотистую кислоту. 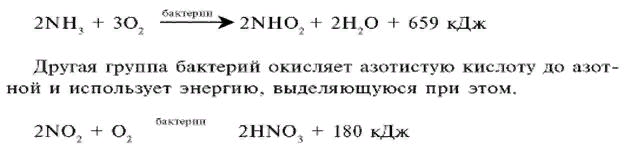 Хемосинтезирующие бактерии играют важную роль в круговороте веществ в природе.  Гетеротрофная ассимиляция Гетеротрофные организмы строят органические вещества своего тела из уже имеющихся готовых органических веществ. К гетеротрофам относят животных, грибы, некоторых бактерий. Гетеротрофные организмы способны строить свои специфические белки, жиры, углеводы только из белков, жиров, углеводов, которые они получают с пищей. В процессе пищеварения эти вещества распадаются до мономеров. Из мономеров в клетках синтезируются вещества, характерные для данного организма. Все эти реакции идут при участии ферментов и с использованием энергии АТФ. 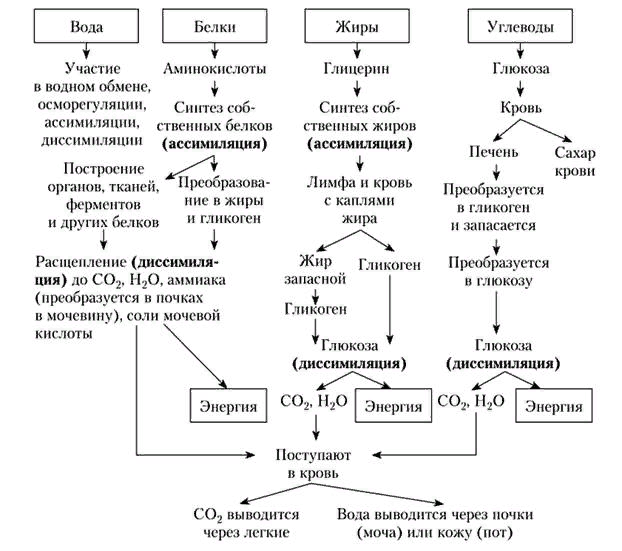 6. Фотосинтез Зеленые растения - фототрофы. Для ассимиляции они используют энергию, выделяемую при окислении неорганических веществ. Зеленые растения имеют в хлоропластах хлорофилл. При участии хлорофилла происходит фотосинтез. Фотосинтез - процесс преобразования солнечной энергии в потенциальную энергию химических связей в органических веществах. Фотосинтез состоит из двух фаз: световой и темновой. Световая фаза. Под действием света молекула хлорофилла, находящаяся в гранах хлоропласта, получает избыток энергии. Часть этой энергии идет на расщепление (фотолиз) молекулы воды. Ионы водорода присоединяют к себе электрон, превращаются в свободный атом водорода. Водород Н идет на восстановление белка - переносчика НАДФ+ (никотинамидадениндинуклеотид фосфат). НАДФ+ + Н НАДФ*H НАДФ*Н переходит в строму хлоропласта, где участвует в синтезе углеводов, в частности, глюкозы. Ионы ОН-, отдав электрон, превращаются в свободные радикалы, которые взаимодействуют друг с другом, образуют воду и свободный кислород. Другая часть энергии используется для синтеза АТФ из АДФ. Таким образом, в световую фазу фотосинтеза образуются: 1) богатое энергетическими связями вещество - АТФ; 2) свободный кислород - О2; 3) происходит присоединение Н (водорода) к переносчику, образуется НАДФ*Н. Реакции световой фазы идут без участия ферментов. Темновая фаза. В темновой фазе происходит связывание СО2. В реакциях темновой фазы участвуют молекулы АТФ и атомы водорода, образовавшиеся в процессе фотолиза и связанные с молекулами-переносчиками. Реакции этой фазы происходят в строме хлоропластов при участии ферментов (Цикл Кальвина). Полученные в результате темновой фазы фотосинтеза молекулы моносахарида - глюкозы через ряд ферментативных реакций превращаются в полисахариды. Так энергия солнечного света преобразуется в энергию химических связей сложных органических веществ. Суммарная реакция фотосинтеза: В результате фотосинтеза образуются органические вещества и кислород атмосферы. 7. Хемосинтез Синтез органических веществ у автотрофных бактерий идет с использованием энергии, выделяющейся при химических реакциях окисления неорганических соединений: сероводорода, серы, аммиака, азотистой кислоты. Источник водорода для восстановления СО2 – вода. Этот процесс называется хемосинтезом. К группе автотрофов-хемосинтетиков относят нитрифицирующие бактерии. Одна группа бактерий получает энергию, необходимую для синтеза органических веществ, в результате реакции окисления аммиака в азотистую кислоту. Хемосинтезирующие бактерии играют важную роль в круговороте веществ в природе. 8. Биосинтез белков в клетках представляет собой последовательность реакций матричного типа, в ходе которых последовательная передача наследственной информации с одного типа молекул на другой приводит к образованию полипептидов с генетически обусловленной структурой. Биосинтез белков представляет собой начальный этап реализации, или экспрессии генетической информации. К главным матричным процессам, обеспечивающим биосинтез белков, относятся транскрипция ДНК и трансляция мРНК. Транскрипция ДНК заключается в переписывании информации с ДНК на мРНК (матричную, или информационную РНК). Трансляция мРНК заключается в переносе информации с мРНК на полипептид. Последовательность матричных реакций при биосинтезе белков можно представить в виде схемы.
На схеме видно, что генетическая информация о структуре белка хранится в виде последовательности триплетов ДНК. При этом лишь одна из цепей ДНК служит матрицей для транскрипции (такая цепь называется транскрибируемой). Вторая цепь является комплементарной по отношению к транскрибируемой и не участвует в синтезе мРНК. Молекула мРНК служит матрицей для синтеза полипептида на рибосомах. Триплеты мРНК, кодирующие определенную аминокислоту, называются кодоны. В трансляции принимают участие молекулы тРНК. Каждая молекул тРНК содержит антикодон– распознающий триплет, в котором последовательность нуклеотидов комплементарна по отношению к определенному кодону мРНК. Каждая молекула тРНК способна переносить строго определенную аминокислоту. Соединение тРНК с аминокислотой называется аминоацил–тРНК. Молекула тРНК по общей конформации напоминает клеверный лист на черешке. «Вершина листа» несет антикодон. Существует 61 тип тРНК с разными антикодонами. К «черешку листа» присоединяется аминокислота (существует 20 аминокислот, участвующих в синтезе полипептида на рибосомах). Каждой молекуле тРНК с определенным антикодоном соответствует строго определенная аминокислота. В то же время, определенной аминокислоте обычно соответствует несколько типов тРНК с разными антикодонами. Аминокислота ковалентно присоединяется к тРНК с помощью ферментов – аминоацил-тРНК-синтетаз. Эта реакция называется аминоацилированием тРНК. На рибосомах к определенному кодону мРНК с помощью специфического белка присоединяется антикодон соответствующей молекулы аминоацил-тРНК. Такое связывание мРНК и аминоацил-тРНК называется кодонзависимым. На рибосомах аминокислоты соединяются между собой с помощью пептидных связей, а освободившиеся молекулы тРНК уходят на поиски свободных аминокислот. Рассмотрим подробнее основные этапы биосинтеза белков. 1 этап.Транскрипция ДНК. На транскрибируемой цепи ДНК с помощью ДНК-зависимой РНК-полимеразы достраивается комплементарная цепь мРНК. Молекула мРНК является точной копией нетранскрибируемой цепи ДНК с той разницей, что вместо дезоксирибонуклеотидов в ее состав входят рибонуклеотиды, в состав которых вместо тимина входит урацил. 2 этап.Процессинг (созревание) мРНК. Синтезированная молекула мРНК (первичный транскрипт) подвергается дополнительным превращениям. В большинстве случаев исходная молекула мРНК разрезается на отдельные фрагменты. Одни фрагменты –интроны– расщепляются до нуклеотидов, а другие –экзоны– сшиваются в зрелую мРНК. Процесс соединения экзонов «без узелков» называетсяс плайсинг. Сплайсинг характерен для эукариот и архебактерий, но иногда встречается и у прокариот. Существует несколько видов сплайсинга. Сущность альтернативного сплайсинга заключается в том, что одни и те же участки исходной мРНК могут быть и интронами, и экзонами. Тогда одному и тому же участку ДНК соответствует несколько типов зрелой мРНК и, соответственно, несколько разных форм одного и того же белка. Сущность транс–сплайсинга заключается в соединение экзонов, кодируемых разными генами (иногда даже из разных хромосом), в одну зрелую молекулу мРНК. 3 этап.Трансляция мРНК. Трансляция (как и все матричные процессы) включает три стадии:инициацию (начало), элонгацию (продолжение), итерминацию (окончание). Инициация. Сущность инициации заключается в образовании пептидной связи между двумя первыми аминокислотами полипептида. Первоначально образуется инициирующий комплекс, в состав которого входят: малая субъединица рибосомы, специфические белки (факторы инициации) и специальная инициаторная метиониновая тРНК с аминокислотой метионином – Мет–тРНКМет. Инициирующий комплекс узнает начало мРНК, присоединяется к ней и скользит до точки инициации (начала) биосинтеза белка: в большинстве случаев это стартовый кодон АУГ. Между стартовым кодоном мРНК и антикодоном метиониновой тРНК происходит кодонзависимое связывание с образованием водородных связей. Затем происходит присоединение большой субъединицы рибосомы. При объединении субъединиц образуется целостная рибосома, которая несет два активных центра (сайта): А–участок (аминоацильный, который служит для присоединения аминоацил-тРНК) и Р–участок (пептидилтрансферазный, который служит для образования пептидной связи между аминокислотами). Первоначально Мет–тРНКМетнаходится на А–участке, но затем перемещается на Р–участок. На освободившийся А–участок поступает аминоацил-тРНК с антикодоном, который комплементарен кодону мРНК, следующему за кодоном АУГ. В нашем примере это Гли–тРНКГлис антикодоном ЦЦГ, который комплементарен кодону ГГЦ. В результате кодонзависимого связывания между кодоном мРНК и антикодоном аминоацил-тРНК образуются водородные связи. Таким образом, на рибосоме рядом оказываются две аминокислоты, между которыми образуется пептидная связь. Ковалентная связь между первой аминокислотой (метионином) и её тРНК разрывается. После образования пептидной связи между двумя первыми аминокислотами рибосома сдвигается на один триплет. В результате происходит транслокация (перемещение) инициаторной метиониновой тРНКМетза пределы рибосомы. Водородная связь между стартовым кодоном и антикодоном инициаторной тРНК разрывается. В результате свободная тРНКМетотщепляется и уходит на поиск своей аминокислоты. Вторая тРНК вместе с аминокислотой (в нашем примере Гли–тРНКГли) в результате транслокации оказывается наР–участке, аА–участок освобождается. Элонгация. Сущность элонгации заключается в присоединении последующих аминокислот, то есть в наращивании полипептидной цепи. Рабочий цикл рибосомы в процессе элонгации состоит из трех шагов: кодонзависимого связывания мРНК и аминоацил-тРНК на А–участке, образования пептидной связи между аминокислотой и растущей полипептидной цепью и транслокации с освобождением А–участка. На освободившийся А–участок поступает аминоацил-тРНК с антикодоном, соответствующим следующему кодону мРНК (в нашем примере это Тир–тРНКТирс антикодоном АУА, который комплементарен кодону УАУ). На рибосоме рядом оказываются две аминокислоты, между которыми образуется пептидная связь. Связь между предыдущей аминокислотой и её тРНК (в нашем примере между глицином и тРНКГли) разрывается. Затем рибосома смещается еще на один триплет, и в результате транслокации тРНК, которая была на Р–участке (в нашем примере тРНКГли), оказывается за пределами рибосомы и отщепляется от мРНК. А–участок освобождается, и рабочий цикл рибосомы начинается сначала. Терминация. Сущность терминации заключается в окончании синтеза полипептидной цепи. В конце концов, рибосома достигает такого кодона мРНК, которому не соответствует ни одна тРНК (и ни одна аминокислота). Существует три таких нонсенс–кодона: УАА («охра»), УАГ («янтарь»), УГА («опал»). На этих кодонах мРНК рабочий цикл рибосомы прерывается, и наращивание полипептида прекращается. Рибосома под воздействием определенных белков вновь разделяется на субъединицы. Как правило, синтезированный полипептид подвергается дальнейшим химическим превращениям. Исходная молекула может разрезаться на отдельные фрагменты; затем одни фрагменты сшиваются, другие гидролизуются до аминокислот. Простые белки могут соединяться с самыми разнообразными веществами, образуя гликопротеины, липопротеины, металлопротеины, хромопротеины и другие сложные белки. Кроме того, аминокислоты уже в составе полипептида могут подвергаться химическим превращениям. Например, аминокислота пролин, входящая в состав белка проколлагена, окисляется до гидроксипролина. В результате из проколлагена образуется коллаген– основной белковый компонент соединительной ткани. Реакции модификации белков не являются реакциями матричного типа. Такие биохимические реакции называются ступенчатыми. Энергетика биосинтеза белков. Биосинтез белков – очень энергоемкий процесс. При аминоацилировании тРНК затрачивается энергия одной связи молекулы АТФ, при кодонзависимом связывании аминоацил-тРНК – энергия одной связи молекулы ГТФ, при перемещении рибосомы на один триплет – энергия одной связи еще одной молекулы ГТФ. В итоге на присоединение аминокислоты к полипептидной цепи затрачивается около 90 кДж/моль. При гидролизе же пептидной связи высвобождается лишь 2 кДж/моль. Таким образом, при биосинтезе большая часть энергии безвозвратно теряется (рассеивается в виде тепла). |