1. Место вирусологии в ряду классических наук. Содержание фундаментальных и прикладных разделов вирусологии, общей и частной вирусологии. Связь вирусологии с другими областями знаний
 Скачать 345.58 Kb. Скачать 345.58 Kb.
|

|
10. Какие вещества кроме белка и НК входят в состав вирусов. Их функции.+ 6вопр Помимо белков в липопротеидной оболочке обнаружены липиды и углеводы. Углеводы преимущественно содержаться в гликопротеидных пепломерах на поверхности вирусной частицы.Выросты, состоящие из липо–или гликопротеидов, выполняют рецепторную функцию. Т.е. у вирионов сложноорганизованных вирусов имеется еще поверхностная оболочка - суперкапсид, содержащий, кроме белков, также углеводы, липиды, компоненты клетки хозяина.Эти рецепторы впоследствии помогают вирусу присоединиться к клетке хозяина и потом проникнуть в нее. А липопротеидная оболочка вируса сливается с мембраной клетки, тем самым обеспечивая прохождения НК вируса в клетку.В составе вирусов обнаружены минеральные вещества К, Na, Ca, Mg, Fe. Они участвуют в формировании связей белка с нуклеиновой кислотой. 11. Что такое рецепторы и антирецепторы. Их состав. Их функция и на каком принципе она основана. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды.Структура рецепторов почти у всех одинакова. Структурно рецептор состоит из трех частей:1)внемембранного, осуществляющего взаимодействие с веществом (лигандом);2) трансмембранного, осуществляющего перенос сигнала;3) цитоплазматического.Эти рецепторы впоследствии помогают вирусу присоединиться к клетке хозяина и проникнуть в нее. Рецепторы распределены в мембране клетки диффузно. Вирусные антирецепторы — это белки и гликопротеины, их также называют прикрепительными белками. Часто они образуют выступы на поверхности вириона (шипы, пепломеры), однако собственно антирецептор (функциональная часть молекулы прикрепительного белка) защищен от случайных взаимодействий и находится в углублении. Взаимодействие между рецептором и антирецептором вируса происходит по принципу комплементарности (дополнительность (пространственная, по заряду, электронной структуре)-основа взаимного сродства молекул). 12. Способы проникновения вирусов в клетку. Механизм реализации проникновения. Термином "вход" обозначается четыре этапа инициации жизненного цикла вирусов в клетках-мишенях. 1эт.Присоединение вирионов к рецепторам клеточной мембраны, при котором происходят первоначальные конформационные изменения белков наружной оболочки вирусов. 2эт. - взаимодействие с корецепторами посредниками проникновения вируса в клетку. Этот процесс сопровождается дальнейшими конформационными изменениями белковых компонентов вирусной оболочки, а также изменениями клеточной плазмалеммы. 3эт.- непосредственное перемещение вируса в клеточной мембране(проникновение) и осуществляемое с помощью различных механизмов. 4эт- освобождение генома вируса от нуклеокапсидной оболочки и начало транскрипции его РНК. Способы проникновения: макропиноцитоз, три вида эндоцитоза (клатрин-зависимый, клатрин-независимый и холестерол-зависимый), с помощью образования кавеол (вогнутости плазматической мембраны) и подобный последнему механизм, зависимый от динамина.1. Вирусные оболочки могут сливаться с клеточной мембраной ( вирус гриппа).2.Вирус попадает в клетку путем пиноцитоза. При этом ферменты клетки хозяина расщепляют его оболочку и высвобождают нуклеиновую кислоту (например, вирус полиомиелита животных).3. Через поврежденные участки клеточной стенки растительных клеток. Потом переходят от одной клетки к другой по цитоплазматическим мостикам. Взаимодействие вируса и клетки может происходить двумя путями: неспецифическим (при случайном столкновении, когда вирионы удерживаются на клеточной поверхности за счет электростатических сил) и специфическим (осуществляется при наличии на клеточной поверхности рецепторов, обеспечивающих прочное прикрепление вируса к клетке). После адгезии вирус проникает через плазмалемму и затем происходит перемещение его генома к определенным участкам цитоплазмы и ядра клетки, где инициируется процесс репродукции. 13. Этапы взаимодействия вируса с клеткой. На каком этапе реализуется генетическая информация вируса. 1.Адсорбция - пусковой механизм, связанный со взаимодействием специфических рецепторов вируса и хозяина. Эффективность проникновения вируса связана большим количеством рецепторов. 2.Проникновение - путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза). 3.Освобождение нуклеиновых кислот - “раздевание” нуклеокапсида и активация нуклеиновой кислоты. 4.Реализация генетической информации. Т. е. синтез нуклеиновых кислот и вирусных белков - подчинение систем клетки хозяина и их работа на воспроизводство вируса. 5.Морфогенез вирусных частиц. Ассоциация реплицированных копий вирусной нуклеиновой кислоты с капсидным белком. 6.Выход вирусных частиц из клетки, приобретения суперкапсида оболочечными вирусами. Ограничения, накладываемые клеткой на вирус. 1. В клетках про- и эукариот (кроме клеток растений) отсутствуют ферменты транскрипции-репликации вирусного РНК-генома. Вирус должен иметь свои ферменты в составе вириона или в закодированном виде в геноме. 2. В цитоплазме клеток нет ферментов для транскрипции ДНК вируса. Следовательно, клеточную ДНК-зависимую РНК-полимеразу могут использовать только ядерные ДНК-содержащие вирусы. 3. В клетках эукариот белоксинтезирующий аппарат приспособлен для трансляции моноцистронных РНК (не распознает внутренние участки инициации). Вирусы должны синтезировать или отдельные мРНК для каждого гена, или мРНК нескольких генов и кодирующий полипротеин, разрезаемый на отдельные белки. 4. В клетках прокариот возможна множественная внутренняя инициация трансляции на полицистронных матрицах. Для вирусов прокариот ограничение 3 снимается.Выработка интерферона,Ликополисахариды,липопротеиды нейтрализуют антирецепторы. 14. Супероболочка вирусов. Её происхождение, этапы формирования. Нуклеоид вирусов покрыт капсидом. Он состоит из капсомеров. А они в свою очередь из белковых субъединиц-протомеров. Капсомеры состоят в тесной связи с нуклеиновой кислотой, поэтому их вместе называют нуклеокапсидом. Капсид защищает НК, способствует переходу ее от одной клетки в другую. Вирусы, которые имеют только нуклеокапсид – простоорганизованные. У некоторых вирусов вирион имеет еще и дополнительную оболочку суперкапсид (пеплос). Такие вирусы- сложноорганизованные. По строению суперкапсид похож на цитоплазматическую мембрану клеток, что обусловлено его клеточным происхождением, так как он формируется в момент выхода вириона из пораженной клетки. Суперкапсид содержит в своем составе углеводы и липиды, а также небольшое количество белков, которые выполняют адресную функцию, то есть обеспечивают прикрепление вириона к поверхности клетки-хозяина. У большинства вирусов гликопротеиды формируют «шипы» на поверхности вирусной частицы. Шипы представляют собой морфологические субъединицы, построенные из нескольких молекул одного и того же белка. Вирусы гриппа имеют два типа шиповУ некоторых вирусов имеется также промежуточная оболочка – белковая мембрана. Этапы:1.Синтезируются глико/липопротеиновые матричные белки.2.Встраиваются с внутренней стороны мембраны. 3Вирусный нуклеопротеид приближается к мембране.4Она его одевает, смыкается5Вирус отпочковывается во внешнюю среду. Особенности выхода из клетки оболочечных и безоболочечных вирусов. Различают два основных типа выхода вирусного потомства из клетки. Первый тип — взрывной — характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имеющих суперкапсидной оболочки. Второй тип — почкование. Он присущ вирусам, имеющим суперкапсидную оболочку. На заключительном этапе сборки нуклеокапсиды сложно устроенных вирусов фиксируются на клеточной плазматической мембране, модифицированной вирусными белками, и постепенно выпячивают ее. В результате выпячивания образуется «почка», содержащая нуклеокапсид. Затем «почка» отделяется от клетки. Таким образом, внешняя оболочка этих вирусов формируется в процессе их выхода из клетки. При таком механизме клетка может продолжительное время продуцировать вирус, сохраняя в той или иной мере свои основные функции. 15. Морфогенез безоболочечных вирусов со спиральным типом симметрии (на примере ВТМ). 1. Образование структуры, состоящей из 2-х дисков, сформированных одинаковыми субъединицами. 2. Образование инициаторной петли РНК. 3. Встранивание петли РНК в отверстие диска (инициация сборки). 4. Элонгация (собственно сборка). Идет в 2-х направлениях вдоль цепи РНК, соповождается закручиванием спирали. 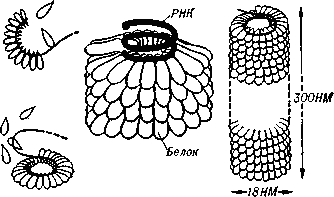 16. Морфогенез безоболочечных вирусов с изометрическим типом симметрии (на примере полиовируса). Все безоболочечные вирусы (Клеточные паразиты, центральный Капсид которых не окружен вирусными оболочками) позвоночных имеют кубическую симметрию. Структурные белки просто устроенных вирусов связываются спонтанно, образуя капсомеры, которые благодаря самосборке образуют капсиды, в которые упаковывается вирусная нуклеиновая кислота. Капсид полиовириона содержит 60 структурных единиц, состоящих из полипептидов VP1, 2, 3, 4. 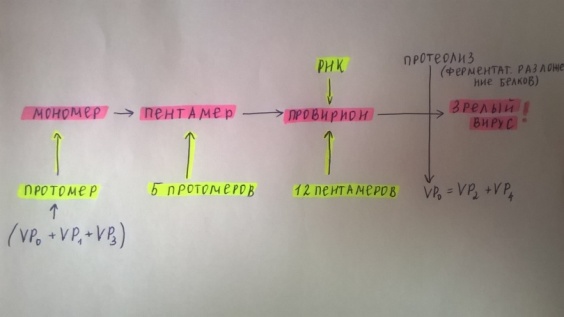 Сборка вирионов с икосаэдральным типом симметрии также протекает по принципу самосборки, однако, как правило, капсид собирается из преформированных структурных единиц.В процессе морфогенеза на первой стадии происходит ассоциация VP0, VP1 и VP3, в результате чего образуется протомер. Пять протомеров агрегируют с образованием пентамера, пентамеры образуют рыхлый провирион. На последней стадии происходит протеолиз VP0 с образованием VP2 и VP4, что приводит к конформационным перестройкам в капсиде и, как следствие, к уплотнению структуры капсида. Сборка вирионов с икосаэдральным типом симметрии также протекает по принципу самосборки, однако, как правило, капсид собирается из преформированных структурных единиц.В процессе морфогенеза на первой стадии происходит ассоциация VP0, VP1 и VP3, в результате чего образуется протомер. Пять протомеров агрегируют с образованием пентамера, пентамеры образуют рыхлый провирион. На последней стадии происходит протеолиз VP0 с образованием VP2 и VP4, что приводит к конформационным перестройкам в капсиде и, как следствие, к уплотнению структуры капсида.17. Формирование супероболочки вирусов. Этапы формирования.
18 Типы взаимодействия вирусов с клеткой в зависимости от исхода инфекции. Подробная характеристика каждого типа взаимодействия. В зависимости от исходов выделяют три типа взаимодействия вируса с клеткой. 1. В клетке образуется новое полноценное поколение вирусных частиц вирионов, а сама клетка погибает вследствие действия вируса(продуктивный тип).Механизмы:цитолиз, образование симпластов и синцитиев, апоптоз зависят от самого вируса. Цитолиз может быть связан с повышением проницаемости клеточной мембраны. Поступление Са в клетку ведет к ионному дисбалансу, вхождению воды в клетку, ее разбуханию, разрыву мембраны и вытеканию цитоплазмы.При множественном почковании оболочечных вирусов возможен феномен «решета», когда клетка не успевает латать бреши и погибает.Аутолизис, связанный с повреждением лизосом.Еще один механизм, приводящий к гибели вирусинфицированные клетки-это формирование под действием вирусов многоядерных клеточных образований (симпластов и синцитиев), не способных выполнять обычные клеточные функции и, вследствие этого, обреченных на гибель.Вирусинфицированные клетки могут гибнуть по механизму апоптоза-запрограммированной клеточной гибели. 2. Биосинтез вируса в клетке не достигает завершения, гибнет вирус, а не клетка, которая возвращается к нормальному функционированию. Такой процесс получил название абортивного. 3. Еще одним типом взаимодействия вируса с клеткой является ее трансформация.Онкогенная трансформация блокирует механизмы гибели клетки путем апоптоза, и клетка приобретает способность к непрерывному росту и делению, что может привести к развитию опухоли. 19. Типы взаимодействия вирусов с клеткой в зависимости от продолжительности пребывания вируса в организме. Подробная характеристика каждого типа взаимодействия. В зависимости от продолжительности пребывания вируса в организме выделяют два главных типа взаимодействия возбудителя с организмом хозяина. Первый тип взаимодействия характеризуется непродолжительным пребыванием вируса в организме, и проявляется в двух формах инфекционного процесса. 1. Острая или продуктивная инфекция. 2. Инапарантная (субклиническая) форма инфекции — бессимптомная инфекция с непродолжительным пребыванием вируса в организме и его выделением во внешнюю среду (например, гепатит A, ротавирусная инфекция). Второй тип взаимодействия характеризуется длительным пребыванием вируса в организме, когда вирус сосуществует с клеткой и может репродуцироваться, не нанося клетке видимого вреда. Такое состояние длительного сосуществования получило название персистентенции. Персистентная инфекция у человека может проявляться в разных формах. 1. Латентная инфекция — при которой вирус «молчит». Инфекционный вирус, находящийся в организме в состоянии провируса (интегрированного в геном клетки хозяина), в нормальных условиях не проявляет себя и не выделяется из клеток организма. Однако в определенных условиях (при снижении иммунитета), он может реактивироваться, вызывая характерные клинические проявления инфекции (например, герпесвирусные инфекции). 2. Хроническая инфекция — вирус присутствует в организме постоянно. Возможно полное отсутствие клинических проявлений инфекции — непрерывный вариант хронической инфекции, и обострение патологического процесса — рецидивирующий вариант хронической инфекции (например, вирусный гепатит C). 3. Медленная инфекция — медленно прогрессирующее заболевание с исключительно длинным латентным периодом (например, ВИЧ-инфекция, переходящая в СПИД). 20 Взаимодействие с клеткой умеренных фагов (этапы взаимодействия). Механизм интеграции генома фага в геном бактериальной клетки. Индукция генома профага. Механизмы индукции. По характеру взаимодействия бактериофага с бактериальной клеткой различают вирулентные и умеренные фаги. Процесс взаимодействия вирулентного бактериофага с кл. включает: адсорбция бактериофага на клетке, проникновение, биосинтез компонентов фага и их сборка, выход бактериофагов из клетки. Первоначально бактериофаги прикрепляются к рецепторам на поверхности бактериальной клетки. Хвост фага с помощью ферментов, растворяет оболочку клетки, сокращается и ДНК инъецируется в клетку.Эта ДНК вызывает полную перестройку метаболизма клетки. ДНК бактериофага начинает транскрибироваться с помощью собственного фермента транскриптазы. Синтезируются иРНК, которые поступают на рибосомы клетки-хозяина. Репликация ДНК бактериофага происходит с участием собственных ДНК-полимераз. После синтеза белков и репликации ДНК наступает созревание и образование зрелых фаговых частиц. Затем происходит лизис клетки, и освобождаются новые зрелые бактериофаги. Также фаг может инициировать лизогенный цикл, при котором он вместо репликации обратимо взаимодействует с генетической системой клетки-хозяина, встриваясь в хромосому. В таком случае гено м фага называют профаг. Профаг, ставший частью хромосо мы клетки, при ее размножении реплицируется синхронно с геномом бактерии, не вызывая ее лизиса, и передается по наслед ству. Бактерия, содержащая профаг, становится лизогенной до тех пор, пока профаг не будет стимулирован на осуществление лизирующего цикла репликации. Переход от лизогении к лизису - индукция профага. На индукцию фага оказывает сильное воздействие состояние клетки хозяина,наличие питательных веществ и др. Скудные условия для роста способствуют лизогенному пути, а хорошие -лизирующему. 21. Что такое лизогенная конверсия. Примеры. Лизогенная или фаговая конверсия (в микробиологии) — изменение свойств бактериальной клетки вследствие встраивания части генома умеренного бактериофага (профаг).Данное явление качественно изменяет свойства бактериальной клетки. Хорошо известны примеры л.к.,когда приобритение профага обеспечивает бакт.кл. доп.фаторами вирулентности,профаговые гены учавствуют в адаптации клетки. Иными словами лизогенная бактерия будет вирулентной, тогда как её нелизогенный двойник останется безвредным. У других бактерий присутствие профага вызывает изменение морфологии или антигенных свойств. Такое изменение генетических свойств, вызванное вирусной ДНК, обозначают терминами «инфекционная наследственность» или «лизогенная (фаговая) конверсия». Примером Ф.к. является приобретение иммунитета к повторному заражению гомологичным фагом.Фаг лямбда. 22. Что такое опухолевая трансформация клетки. Какие РНК-содержащие вирусы вызывают опухолевую трансформацию. К какой группе вирусов относится эта группа онкогенных РНК-содержащих вирусов. Механизм опухолевой трансформации онкогенными РНК-содержащими вирусами. ОПУХОЛЕВАЯ ТРАНСФОРМАЦИЯ, критический этап онкогенеза - момент превращения нормальной клетки в опухолевую.В основе превращения нормальной клетки в злокачественную – опухолевая трансформация – лежит перенос или иное взаимодействие ДНК. Агент, вызывающий пролиферацию клеток, - это продукт гена.(в.лейкомы роговицы,лейкоза крупного рогатого скота,опухоли молочной железы мышей)Онкогенез, вызываемый у животных РНК-вирусами. К образованию опухолей у животных могут быть причастны и РНК-вирусы – ретровирусы. Они относятся к икосаэдриеским вирусам с оболочкой и содержат (+)РНК-геном (одноцепочную РНК). Вызывают саркому Рауса у кур и лейкемию у мышей. Название «ретровирусы» связано с тем, что в их размножении участвует обратная транскриптаза. РНК этих вирусов не может воспроизводиться путем простой репликации – необходима ее предварительная транскрипция в ДНК с последующей интеграцией этой ДНК в одну из хромосом клетки-хозяина. Интеграция – необходимый этап репродукции вируса; только интегрированная вирусная ДНК будет транскрибироваться. Т.к. интеграция в клеточную ДНК входит в жизненный цикл вируса, частота интеграции очень высока. Вероятно, вируснаяДНКможет включаться в клеточную в любом месте. Размножение вируса не приводит к лизису клетки. Нуклеокапсид образуется внутри клетки, перемещается затем к плазматической мембране и выходит наружу, одетый в оболочку из этой мембраны. Интегрированная ДНК ретровируса реплицируется вместе с геномом клетки-хозяина и поэтому содержится в каждой клетке опухоли (саркомы). Опухолевый рост клеток обусловлен вирусным геном «src». Этот ген кодирует белок, который представляет собой киназу, фосфорилирующую белки. Эта киназа участвует в преобразовании дифференцированной клетки в клетку эмбрионального типа. Src – ген животного происхождения, который в результате неточной транскрипции был включен в РНК вируса вместе с вирусными генами и закрепился в ней. Таким образом, опухоли, вызываемые ретровирусами, в конечном счете обусловлены переносом какого-то гена животного происхождения в клетку животного. |