Гиста 1 итоговая. гиста итог 1. 1. Общая характеристика и классификация эндокринных желез. Понятие о гормонах, клеткахмишенях и их рецепторах
 Скачать 79.36 Kb. Скачать 79.36 Kb.
|

|
1. Общая характеристика и классификация эндокринных желез. Понятие о гормонах, клетках-мишенях и их рецепторах. Эндокринными называют железы, продукты которых (гормоны) выделяются непосредственно в кровь. Железы могут быть: одноклеточными (элементы ДЭС), в виде мелких компактных клеточных скоплений (панкреатические островки), крупными органными структурами. Некоторые эндокринные железы имеют в составе ткани, имеющие различное эмбриональное происхождение, некоторые могут периодически появляться и исчезать (желтое тело и фолликул яичника) Органные эндокринные железы – покрыты капсулой (ПСТ) с трабекулами (РСТ), несущими сосуды и нервы. Клетки желез образуют тяжи и тесно прилежат к капиллярам, Капилляры желез – фенестрированные или синусоидные (повышенная проницаемость), располагаются очень густыми сетями. Клетки эндокринных желез имеют высокую секреторную активность, их строение зависит от природы вырабатываемого гормона. 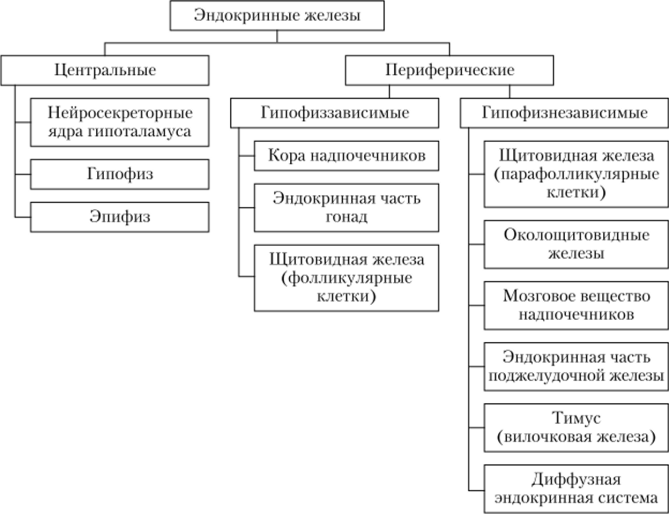 Гормоны – вещества с высокой биологической активностью – регулируют рост и деятельность клеток различных тканей (клеток-мишеней) благодаря наличию на них специфических рецепторов гормонов. 2. Щитовидная железа. Источники развития. Общая морфофункциональная характеристика. Фолликулы, строение стенки фолликулов. Коллоид. Перестройка фолликулов в связи с различной функциональной активностью. Щитовидная железа содержит 2 клеточных дифферона эндокриноцитов, развивающихся из стволовых клеток различного происхождения: T-тироциты (фолликулярные клетки, наиболее многочисленные, вырабатывающие тироксин и трийодтиронин), и С-тироциты (парафолликулярные клетки, вырабатывающие тирокальцитонин) Зачаток щитовидной железы возникает на 3-4 неделе как выпячивание стенки глотки между I и II парами жаберных карманов, которое растет вдоль глоточной кишки в виде эпителиального тяжа. На уровне III-IV пары жаберных карманов тяж раздваивается, дает начало правой и левой долям железы. Начальный тяж атрофируется и дает перешеек, связывающий доли. Зачатки долей быстро разрастаются, образуя рыхлые сети ветевящихся эпителиальных трабекул, клетки которых дифференцируются в Т-тироциты, которые объединяются в фолликулы, между которыми врастает мезенхима с сосудами и нервами. В зачаток железы также врастают производные V пары жаберных карманов – С-тироциты (нейроэктодермальная природа). Строение. Щитовидная железа окружена капсулой, внутрь отходят трабекулы, делящие железу на дольки, в которых располагаются сосуды МЦР и нервы. Основные структурные компоненты паренхимы железы – фолликулы – замкнутые шаровидные или слегка вытянутые образования разных размеров с полостью внутри, выстланные одним слоем эпителиоцитов, представленных Т-тироцитами (фолликулярными клетками), а также С-тироцитами (парафолликулярными клетками нейрального происхождения). В дольке железы различают фолликулярные комплексы (микродольки), которые состоят из группы фолликулов, окруженных тонкой капсулой. В просвете фолликулов накапливается коллоид – секреторный продукт Т-тироцитов, представляющий собой вязкую жидкость, состоящую из тироглобулина. Размер фолликулов и Т-тироцитов, образующих их стенку, варьирует (d= 0.02-0.9 мм). В небольших формирующихся фолликулах, еще не заполненных коллоидом, эпителий однослойный призматический. По мере накопления коллоида размеры фолликулов увеличиваются, эпителий – кубический, затем – плоский. Большая часть фолликулов выстлана Т-тироцитами кубической формы. Увеличение размеров фолликулов обусловлено пролиферацией, ростом и дифференцировкой Т-тироцитов с накоплением коллоида в полости фолликула. 3. Секреторный цикл тироцитов. Роль гормонов тироцитов. Различают основные фазы: фазу продукции и фазу секреции гормонов. Фаза продукции включает: 1) поступление предшественников тироглобулина (а/к, углеводов, воды, йодидов), приносимых из КРС в Т-тироциты. 2) синтез полипептидных цепочек тироглобулина в грЭПС и их гликозилирование в КГ. 3) синтез тиропероксидазы, окисляющей йодиды и обеспечивающей их соединение с тироглобулином на поверхности Т-тироцитов. 4)образование коллоида (йодирование тироглобулина). Фаза выведения: 1) эндоцитоз тиреоидных гормонов, которые подвергаются гидролизу с помощью лизосомных мембран. 2) тироглобулин расщепляется до аминокислот, а монойодтирозин, дийодтирозин, трийодтиронин и тетрайодтиронин освобождаются в цилотплазма. 3) Два последних выводятся через базальную мембрану в капилляры и лифматические капилляры. 4) монойодтирозин и дийодтирозин используются для синтеза новой молекулы тироглобулина. Тироксин и трийодтиронин – мощные стимуляторы окислительных процессов в клетках, (Т3 в 5-10 раз активнее Т4). Усиление обмена веществ, синтеза белков, газообмена, обмена углеводов и жиров. Значительное влияние на развитие, рост и дифференцировку клеток и тканей. (ускоряют рост костной ткани, большое влияние на гистогенез нервной ткани). Тиреоидные гормоны оказывают стимулирующее действие на регенерационные процессы в тканях. Тирокальцитонин (без йода) – участвует в регуляции кальциевого и фосфорного обмена. 4. Парафолликулярные эндокриноциты. Источники развития, локализация, строение, гормоны. С-тироциты. Источники развития По мере развития железы в ее зачаток врастаются производные V пары жаберных карманов – клетки ультимобранхиальных телец. Это С-тироциты нейроэктодермальной природы, они внедряются в закладку щитовидной железы через ультимобранхиальные зачатки. Локализация Во взрослом организме С-тироциты локализуются в стенке фолликулов, залегая между основания соседних Т-тироцитов, но не достигают своей верхушкой просвета фолликула (интраэпителиально). Также располагаются в межфолликулярных прослойках соединительной ткани. Строение По размерам крупнее Т-тироцитов, имеют округлую или угловатую форму. В отличие от Т-тироцитов не поглощают йод, но совмещают образование нейроаминов (норадреналина и серотонина) с биосинтезом белковых гормонов – кальцитонина и соматостатина. Секреторные гранулы густо заполняют ЦП С-тироцитов, имеют сильную осмиофилию и аргирофилию. В ЦП хорошо развиты грЭПС и КГ. Секреторные гранулы двух типов. Мелкие, сильно осмиофильные, вырабатывающие кальцитонин. Более крупные, но слабо осмиофильные – вырабатывают соматостатин) 5. Околощитовидные железы. Источники развития. Общая морфофункциональная характеристика. Роль в регуляции кальциевого обмена. Развитие. Закладываются как выступы из эпителия III и IV пар жаберных карманов глоточной кишки. На 5-6 неделе образуются 4 зачатка желез в виде эпителиальных почек. На 7-8 неделе эти почки отшнуровываются от стенок жаберных карманов, присоединяясь к задней поверхности щитовидной железы. В процессе гистогенеза клетки дифференцируются, растут, ЦП становится светлой (главные паратироциты). На 5 месяце дифференцируются на светлые и темные. На 10 году появляются оксифильные паратироциты. Также могут находиться С-клетки, вырабатывающие кальцитонин. Строение Железа окружена капсулой, паренхима представлена трабекулами (тяжами эпителия, либо скоплениями паратироцитов), разделенными прослойками РСТ с многочисленными капиллярами. Связь между клетками: межклеточные щели, интердигитации и десмосомы. Различают главные паратироциты и оксифильные паратироциты. Главные клетки. Секретируют паратирин, преоблают, маленькие (7-10 мкм), полигональной формы. В периферических зонах ЦП базофильна, там рассеяны скопления полисом. Секреторные гранулы d=150-200 нм. При усилении секреции главные клетки увеличиваются. Среди главных различают – светлые (неактивные) и темные (активные – хорошо развиты органеллы синтеза секрета). 1 темный – 3-5 светлых Оксифильные паратироциты. Малочисленны, поодиночке или группами, крупнее главных. В ЦП оксифильные гранулы, много МХ, слабый КГ. Их рассматривают как стареющие формы главных клеток. Роль в регуляции кальциевого обмена. Околощитовидная железа по принципу обратной связи быстро реагирует на малейшие колебания содержания кальция в крови. Ее деятельность усиливается при гипокальциемии и ослабляется при гиперкальциемии. Паратироциты обладают рецепторами, способными непосредственно воспринимать прямые влияния на них ионов кальция. 6. Надпочечники. Источники развития. Общая морфофункциональная характеристика. Зоны коры и их клеточный состав. Роль гормонов коры надпочечников. Надпочечники – парные эндокринные железы, образованные соединением двух самостоятельных желез, составляющих корковое и мозговое вещество (разное происхождение). Источники развития Закладка корковой части на 5 неделе в виде утолщений целомического эпителия по обе стороны корня брыжейки. Далее эти эпителиальные утолщения (обр. крупными клетками с ацидофильной ЦП) собираются в компактное интерреналовое тело. Ацидофильные клетки становятся зачатком первичной коры надпочечников. На 10 неделе первичная кора окружается снаружи мелкими базофильными клетками (из целомического эпителия), которые дают начало дефинитивной коре надпочечников. Мозговая часть закладывается на 6-7 неделе в результате миграции клеток нервного гребня. В процессе миграции происходит дифференцировка клеток в симпатобласты и хромаффинобласты. Последние внедряются в интерреналовое тело, где размножаются, дифференцируются в хромаффиноциты и дают начало мозговому веществу надпочечников. Железистые (хромаффинные) клетки мозгового вещества – нейроэндокринные. Общая морфофункциональная характеристика. Снаружи надпочечник покрыт капсулой, в которой различают наружный (плотный) и внутренний (рыхлый) слои. В толще капсулы иногда есть скопления клеток коры в виде узелков. От капсулы в корковое вещество отходят тонкие трабекулы с сосудами и нервами. От трабекул и капсулы вглубь органа проникают ретикулиновые волокна, образующие тонкую сеточку вокруг клеток паренхимы в корковом и мозговом веществах. Основные структурные компоненты надпочечников – корковое и мозговое вещество. Корковое вещество занимает около 70-80% объема. В корковом веществе – глюкокортикоиды, минералокортикоиды, половые гормоны. Функция коры кроме клубочковой зоны контролируется АКТГ гипофиза и гормонами почек –ренин-ангиотензивной системой. В мозговом веществе – эпинефрин и норэпинефрин. Зоны коры и их клеточный состав. Корковое вещество образовано 3 зонами: (1) клубочковой – тонкой наружной, (2) пучковой – основная масса коры, (3) сетчатой – узкая внутренняя, прилежащая к мозговому веществу. 1. Клубочковая зона образована небольшими клетками с равномерно окрашенной ЦП, которые формируют круглые арки («клубочки»). Хорошо: аЭПС и КГ, МХ – пластинчатые и трабекулярные кристы. Липидных капель меньше, чем в пучковой зоне 2. Пучковая зона образована крупными оксифильными вакуолизированными клетками (спонгиоцитами), которые образуют радиальные тяжи («пучки»), разделенные синусоидными капиллярами. Хорошо: аЭПС, КГ, МХ – тубулярные кристы. Липидных капель много. 3. Сетчатая зона образована анастомозирующими эпителиальными тяжами, между которыми капилляры. Клетки меньше. Хорошо: аЭПС, МХ – тубулярные кристы, лизосомы и липофусциновые гранулы. Липидных капель меньше. Клетки гибнут апоптозом. Роль гормонов коры надпочечников. Синтезируются в клубочковой, пучковой и сетчатой зонах соответственно: 1) Минералокортикоиды – уровень электролитов и АД (АЛЬДОСТЕРОН – увеличивает реабсорбцию натрия, хлора, бикарбоната, усиливает экскрецию калия и водорода.) Усиливают воспалительные процессы. 2) Глюкокортикоиды – различные виды обмена (углеводный особенно), иммунная система. (КОРТИЗОЛ, кортикостерон, кортизон). Форсируют глюконеогенез, отложение гликогена в печени и миокарде. 3) Половые стероиды (ДЕГИДРОЭПИАНДРОСТЕРОН и его сульфат, андрогенстериодный гормон, эстрогены и прогестерон, но мало) Субстрат для синтеза кортикостероидов – холестерин, извлекается из крови путем рецепторно-опосредованного эндоцитоза в составе ЛПНП, отщепляется гидролизом в лизосомах и накапливается в липидных каплях. Стероиды образуются и выделяются непрерывно, ферментные системы стероидогенеза расположены в аЭПС и МХ. Регуляция клеток коры в пучковой и сетчатой зонах – АКТГ (рецепторный мех-м), а клубочковая зона – зависит от ренин-ангиотензивной системы 7. Мозговое вещество надпочечников. Строение, клеточный состав, гормоны. Отделено от коркового вещества тонкой прослойкой соединительной ткани. Образовано скоплением крупных круглых хромаффинных клеток, между которыми находятся синусоиды. Различают светлые хромаффиноциты (адреноциты) и темные хромаффиноциты (норадреноциты). ЦП густо заполнена плотными секреторными гранулами (адреналин, норадреналин, ДОФА – амины, энкефалины и хромогранины – пептиды). В мозговом веществе находятся мультиполярные нейроны автономной НС. Катехоламины оказывают влияние на ГМЦ сосудов, пищеварительного тракта, бронхов, на сердечную мышцу, а также на метаболизм углеводов (глюкогенолиз, глюконеогенез), липидов (липолиз). В состав мозгового вещества также входят ганглиозные (мало, представляют собой вегетативные нейроны) и поддерживающие клетки (отростчатые, глиальной природы, охватывают хромаффиноциты). 8. Поджелудочная железа. Строение эндокринного отдела. Типы инсулоцитов, гормоны и их действие на организм. Поджелудочная железа – железа смешанной секреции, состоит из экзокринной и эндокринной частей. Эндокринная часть – представлена панкреатическими островками Лангерганса, разбросанными по всей железе в виде округлых компактных скоплений инсулоцитов между ацинусами. Общее количество островков – 0.5-2 млн, размеры 100-200мкм, относительный объем 1-2%, большая часть в хвостовой части железы. Панкреатические островки состоят из нескольких сотен или тысяч инсулоцитов, между которыми: ретикулярные волокна фенестрированные кровеносные капилляры и нервные волокна Инсулоциты характеризуются: овальной или полигональной формой, развитым синтетическим аппаратом образованием и накоплением секреторных гранул Выделяют 5 основных типов инсулоцитов (А- , В-, D-, D1- и PP-клетки) А) А-клетки (ацидофильные) (20-25%), по периферии островков. Плотные гранулы с узким светлым ободком содержат глюкагон – гормон, повышающий уровень глюкозы в крови. Б) В-клетки (базофильные) (60-70%), в центре островков. В гранулах центральная плотная часть неправильной формы окружена широким светлым ободком и содержит кристаллоиды (комплекс инсулин+Zn). В) D-клетки (дендритические) (5-10%) по периферии островков. Равномерно заполненные веществом умеренной плотности гранулы, содержащие соматостатин, обладающий многими функциями, в частности угнетающий секрецию А- и В-клеток островков, а также панкреатоцитов. Г) D1-клетки – разновидность D-клеток. Гранулы с однородным матриксом содержат вазоактивный интестинальный пептид (ВИП) – гормон, вызывающий снижение давления, усиление выделения панкреатического сока. Д) РР-клетки (2-5%). По периферии островков, но могут встречаться и в экзокринном отделе. Полиморфные мелкие гранулы с гомогенным матриксом, в которых находится панкреатический пептид – гормон, угнетающий активность ациноцитов поджелудочной железы. Также имеются промежуточные или ациноостровковые клетки, располагающиеся группами вокруг островков. Гранулы двух типов – зимогенные крупные и мелкие эндокринные. МХ делятся на крупные и мелкие, грЭПС занимает по развитию промежуточное положение. Классифицируют по гормональному профилю на А, В и D (т.к. гранулы напоминают гранулы одного из видов клеток островков). 9. Гипоталамус. Нейросекреция. Морфофункциональная характеристики крупноклеточных и мелкоклеточных ядер гипоталамуса, их роль в регуляции эндокринной системы. Гипоталамус – участок промежуточного мозга, содержащий нейросекреторные ядра, клетки которых вырабатывают и секретируют в кровь нейрогормоны. Эти клетки получают импульсы из других отделов мозга, а их аксоны оканчиваются на кровеносных сосудах (аксовазальные синапсы) Нейросекреторные клетки – отростчатые, крупное везикулярное ядро, базофильны, развитая грЭПС и КГ, от которого отделяются нейросекреторные гранулы. Транспорт гранул: 1) По аксону 1-4 мм/ч вдоль центрального пучка нейротрубочек и нейрофиламентов, 2) Местами накапливаются, растягивая аксон и образуя НАКОПИТЕЛЬНЫЕ НЕЙРОСЕКРЕТОРНЫЕ ТЕЛЬЦА ХЕРРИНГА, в них сосредоточено до 60% всего нейросекрета, лишь 30% в области терминалей. Нейросекреторные ядра гипоталамуса (по размеру клеток и их функциям) делят на крупноклеточные и мелкоклеточные. 1. Крупноклеточные ядра – образованы крупными клеточными телами (в 2-3 р. Больше, чем в других отделах гипоталамуса); к ним относятся: супраоптическое (СОЯ) и паравентрикулярное (ПВЯ) ядра. СОЯ в 3-4 раза больше ПВЯ. Аксоны клеток СОЯ и ПВЯ покидают гипоталамус и в составе гипоталамо-гипофизарного тракта пересекая гемато-энцефалический барьер, проникают в заднюю долю гипофиза, где образуют терминали на капиллярах. Крупноклеточные ядра секретируют: 1) АДГ/ВП – образуется в СОЯ – усиление тонуса ГМЦ артериол, приводящее к повышению АД, обеспечивая реабсорбцию 99% жидкости из мочи. 2)Окситоцин – образуется в ПВЯ – обеспечение сокращения мышечной оболочки матки во время родов, сокращение миоэпителиальных клеток в концевых отделах молочной железы, выброс молока в протоки 2. Мелкоклеточные ядра. Вырабатывают ряд гипофизотропных факторов: Рилизинг факторы/либерины – усиливают выработку гормонов клетками передней доли гипофиза, попадая к ним по воротной системе сосудов. (соматолиберин, тиролиберин, кортиколиберин, фоллиберин,, люлиберин, пролактолиберин, меланолиберин) Ингибирующие факторы/статины – уменьшают выработку гормонов клетками передней доли гипофиза, попадая к ним по воротной системе сосудов. (соматостатин, пролактостатин, меланостатин) Аксоны нейросекреторных клеток этих ядер образуют терминали на первичной капиллярной сети в срединном возвышении. Эта сеть дальше собирается в воротные вены, проникающие в переднюю долю гипофиза и распадающиеся на вторичную сеть капилляров между тяжами аденоцитов. 10. Гипофиз. Источники и ход эмбрионального развития гипофиза. Гипофиз регулирует активность ряда желез внутренней секреции и служит местом выделения гипоталамических гормонов крупноклеточных ядер гипоталамуса. Состоит из двух эмбриологически структурно и функционально различных частей – нейрогипофиза – выроста промежуточного мозга и аденогипофиза, ведущей тканью которого служит эпителий. Аденогипофиз разделяется на крупную переднюю долю, узкую промежуточную и слабо развитую туберальную часть. Аденогипофиз развивается из гипофизарного кармана Ратке выстилки верхней части ротовой полости. Аденоциты – эпителиального происхождения. Нейрогипофиз образуется как выпячивание промежуточного пузыря закладки головного мозга. Дивертикул – выпячивание промежуточного мозга. Развитие. Закладка гипофиза на 4-5 неделе, как результат взаимодействия двух отдельных зачатков – эпителиального и нейрального. Из эктодермального эпителия, выстилающего ротовую ямку зародыша, выпячивается гипофизарный карман Ратке, направляющийся к основанию формирующегося головного мозга и дающий начало аденогипофзу. Дифференцировка начинается после его соприкосновения с протовоположным выпячиванием промежуточного пузыря зачатка головного мозга. Базальная часть промежуточного пузыря дает начало гипоталамузу. Дифференцировка кармана Ратке начинается разрастанием его передней стенки, которая станет передней долей гипофиза и туберальной ее частью. Задняя стенка того же кармана станет промежуточной долей. Между передней и промежуточной долями иногда сохраняется остаток полости гипофизарного кармана в виде гипофизарной щели. Нейроглия дистального конца воронки разрастается и формирет заднюю долю гипофиза (нейрогипофиз). Проксимальная часть воронки суживается и становится гипофизарной ножкой (стеблем), связывающей гипоталамус и гипофиз. Происходит дифференцировка эпителиоцитов аденогипофиза и образуются клеточные диффероны хромофильных и хромофобных эндокриноцитов. 11. Общая морфофункциональная характеристика и клеточный состав аденогипофиза. Хромобные и хромофильные аденоциты. Гормоны аденогипофиза, их действие на организм. Аденогипофиз – образован разветвленными эпителиальными тяжами – трабекулами, формирующими густую сеть, между трабекулами РВСТ и синусоидные капилляры. Трабекулы образованы эндокриноцитами. По периферии трабекул – хромофилы. По центру трабекул – хромофобы. Хромофилы: Развитый синтетический аппарат Накопление гранул Бывают ацидофилы и базофилы А) Ацидофилы (40%): Мелкие Круглые Хорошо развиты органеллы Много крупных гранул Делятся на: Соматотропы – СТГ – стимуляция роста Лактотропы – ЛТГ – развитие молочных желез и лактация Б) Базофилы (10-20%): Крупные Мало мелких гранул Включают: Гонадотропы – вырабатывают: а) фолликулостимулирующий Г., б) лютеинизирующий Г. (секреция женских и мужских гормонов, развитие овуляции и формирование желтого тела). Тиротропы – ТТГ Кортикотропы – АКТГ, продукт расщепления проопимеланокортина. Хромофобы: (1) Хромофилы после выведения гранул (2) Камбиальные элементы (3) Фолликулярно-звездчатые клетки – несекреторные, охватывают секреторные клетки, фагоцитируют гибнущие клетки, влияют на секреторную активность базофилов и ацидофилов. Промежуточная доля гипофиза - Развита очень слабо - Состоит из прерывистых тяжей базофильных и хромофобных клеток - Секретируют меланоцитостимулирующий гормон (МСГ, активирует меланоциты) и липотропный гормон (ЛГ, стимулирует обмен жиров) - Встречаются кистозные полости, выстланные реснитчатыми клетками и содержащие коллоид. Туберальная часть гипофиза В виде тонкого рукава покрывает гипофизарную ножку, отделяясь от нее узким слоем соединительной ткани. Она состоит из тяжей хромофобных и хромофильных клеток. 12. Портальная система кровообращения гипофиза, гипоталамо-аденогипофизарная система. Приносящие гипофизарные артерии вступают в медиальное возвышение медиобазального гипоталамуса, где разветвляются в сеть капилляров (первичное капиллярное сплетение портальной системы). Эти капилляры образуют петли и клубочки, с которыми контактируют терминали аксонов нейросекреторных клеток аденогипофизотропной зоны гипоталамуса (мелкоклеточные ядра выделили рилизинг факторы). Далее капилляры первичного сплетения собирают в портальные вены, идущие вдоль гипофизарной ножки в переднюю долю, где они распадаются на синусоидные капилляры (вторичная капиллярная сеть), разветвляющиеся между трабекулами паренхимы железы. Наконец, синусоиды вторичной капиллярной сети собираются в выносящие вены, по которым богатая гормонами передней доли кровь поступает в общую циркуляцию. 13. Строение и функция нейрогипофиза, гипоталамо-нейрогипофизарная система. Задняя доля гипофиза образована клетками эпендимы . Они имеют отростчатую форму и называются ПИТУИЦИТАМИ, их многочисленными тонкие отростки заканчиваются в адвентиции КРС или на БМ капилляров. В задней доле гипофиза накапливаются АДГ и окситоцин, которые вырабатываются крупноклеточными ядрами гипоталамуса. Аксоны крупноклеточных ядер собираются в гипоталамо-нейрогипофизарные пучки, входят в нейрогипофиз, где заканчиваются крупными терминалями (тельцами Херринга). Они формируют нейрососудистые (нейрогемальные) синапсы, посредством которых нейросекрет поступает в кровь. 14. Эпифиз. Источники развития. Общая морфофункциональная характеристика, клеточный состав. Возрастные изменения. Эпифиз – нейроэндокринный орган, получающий информацию из нервной и эндокринной систем, которая интегрируется в нем и регулирует активность его клеток – пинеалоцитов. Регулирует циклические процессы в организме (деятельность репродуктивной системы), циркадные ритмы. Развитие. Развивается как выпячивание крыши III желудочка промежуточного мозга на 5-6 нед. В его состав включается субкомиссуральный орган, который развивается из эпендимы III желудочка. Он сильно редуцирован. В результате дифференцировки нейральных стволовых клеток развиваются два клеточных дифферона – пинеалоцитарный и глиоцитарный. Эпифиз покрыт тонкой капсулой, от которой отходят септы с КРС и нервами, которые разделяют орган на дольки. Паренхима долек состоит из анастомозирующих клеточных тяжей, групп и фолликулов из пинеалоцитов и интерстициальных клеток. Пинеалоциты (90%). Отростчатой формы Округлое ядро с инвагинациями Крупное ядрышко ЦП: Хорошо: грЭПС, аЭПС, КГ, МХ, ЛС, РС В ЦП есть синаптические ленты (пластинки с пузырьками типа синаптических) Связаны ЩК и ДС На концах отростков есть булавовидные расширения с пузырьками, оканчивающиеся на капиллярах. Выделяют светлые и темные пинеалоциты. Вырабатывают два типа веществ: индоламины (мелатонин - антагонист МСГ, угнетает секрецию гонадолиберина, снижая активность гонад) и пептиды (аргинин-вазотоцин – угнетает секрецию ФСГ и ЛГ; пинеальный антигонадотропный пептид - ПАГТП). Интерстициальные клетки (5%) Длинные отростки, окружающие пинеалоциты и проникающие в перикапиллярное пространство Ядро удлиненное, плотное ЦП: умеренные органеллы, толстые пучки филаментов. Являются видоизмененными астроцитами и выполняют опорную функцию Возрастные изменения У человека эпифиз достигает максимального развития к 5-6 годам жизни, после чего, несмотря на продолжающееся функционирование, начинается его возрастная инволюция. Некоторое количество пинеалоцитов претерпевают атрофию, а строма разрастается, и в ней увеличивается отложение фосфатных и карбонатных солей в виде слоистых шариков, называемых мозговым песком (ацервулюс) 15. Тимус. Эмбриональное развитие. Особенности строения эпителиоретикулярной стромы тимуса. Строение и значение гематотимического барьера. Тимус – центральный орган иммунной системы, в котором происходит АНПД Т-лимфоцитов из предшественников, поступающих из ККМ. Развитие. Закладка в конце 1 месяца из ЭПИТЕЛИЯ ГЛОТОЧНОЙ КИШКИ в области III и IV пар жаберных карманов в виде тяжей многослойного эпителия. Дистальная часть зачатков III пары утолщается и образует тело тимуса, а проксимальная – вытягивается. Тимус обособляется от жаберного кармана. Правый и левый зачатки тимуса срастаются. На 7 неделе появляются первые лимфоциты. На 8-11 неделе мезенхима подразделяет закладку тимуса на дольки. 11-12 недели – дифференцировка лимфоцитов. На 3 месяце – образование корковой и мозговой частей. В течение 3-5 месяцев наблюдается дифференцировка клеток и появление различных типов ретикулярных эпителиоцитов. К 6 месяцам развитие заканчивается. Строение стромы Тимус покрыт капсулой, которая продолжается в перегородки с КРС , разделяющие его на дольки. Долька состоит из трехмерной сети отростчатых эпителиальных (эпителиоретикулярных) клеток, образующих строму тимуса, в петлях которой лимфоциты. В каждой дольке выделяют корковое и мозговое вещество. Гематотимический барьер - Имеется только в корковом веществе - Предотвращает воздействие циркулирующих в крови антигенов на созревающие тимоциты В состав барьера входят: Эндотелий капилляра (м/б перициты) БМ капилляра Перикапиллярное пространство (содержит волокна и макрофаги) БМ эпителиоретикулоцитов ЦП эпителиоретикулоцитов (связанных десмосомами) 16. Строение и тканевой состав коркового и мозгового вещества дольки тимуса. Роль тимуса в лимфоцитопоэзе. 1. Корковое вещество – более темное вследствие плотной упаковки тимоцитов. Лимфоцитопоэз: 1) Претимоциты поступают в корковое вещество из ККМ; 2) пролиферирующие тимоциты располагаются скоплениями между эпителиоретикулоцитами в субкапсулярной зоне, они большие и без рецепторов Т-клеток. 3) Созревающие тимоциты продолжают делиться и перемещаются в глубокие части коры, образуют РТК за счет реаранжировки генома. Они имеют вид средних и малых лимфоцитов. 4) Подавляющее большинство тимоцитов образовавшихся в коре гибнет там же апоптозом в процессе отбора, включающего положительную селекцию (выжвание клеток, способных распознать собственные белки MHC) и отрицательную селекцию (гибель клеток с рецепторами к собственным антигенам). Погибшие клетки уничтожаются макрофагами. 5) В коре образуются субпопуляции Т-клеток (сначала экспрессируют CD4+CD8+РТК, затем РТК и только один из маркеров). 6) Более зрелые Т-клетки перемещаются в мозговое вещество. Эпителиоретикулярные клетки – охватывают отростками тимоциты, создавая микроокружение, необходимое для их деления и созрения. Есть несколько особых вариантов эпителиоретикулоцитов: (1) секреторные клетки – содержат гранулы, вырабаывают факторы созревания тимоцитов: тимозин, тимопоэтин, тимусный сывороточный фактор. (2) «клетки-няньки» - заключают в своей ЦП несколько десятков делящихся и гибнущих тимоцитов, изолируя их от окружающих клеток и участвуя в их селекции. (3) периваскулярные клетки – охватывают уплощенные отростками капилляры и служат элементом гемато-тимусного барьера. 2. Мозговое вещество – светлее коркового, меньше более зрелых тимоцитов, которые покидают тимус и заселяют Т-зависимые зоны периферических органов иммунной системы. Эпителиоциты более крупные , их больше. В отдельных участках ороговевают, накладываются друг на друга и образуют слоистые эпителиальные тельца Гассаля. 17. Лимфатические узлы. Общая морфофункциональная характеристика. Характеристика коркового вещества. В- и Т-зависимые зоны. Лимфатические узлы – периферические органы иммунной системы, располагающиеся по ходу лимфатических сосудов. Имеют бобовидную форму, к выпуклой поверхности подходят приносящие лимфатические сосуды, в области ворот на вогнутой поверхности входят артерии и нервы и выходят выносящие лимфатические сосуды и вены. Покрыты капсулой, от которой отходят трабекулы. Строма узлов образована трехмерной сетью ретикулярных клеток, коллагеновых и ретикулярных волокон, а также макрофагами и АПК. В петлях стромы располагаются элементы лимфоцитарного ряда. В каждом узле можно выделить корковое и мозговое вещество. Корковое вещество Наружная кора (под капсулой узла) Глубокая кора (паракортикальная зона) Наружная кора включает: Лимфоидную ткань, образующую лимфатические узелки (В-зависимые зоны) Межузелковые скопления Синусы – под капсулой по ходу трабекул Лимфатический узелок – сферическое скопление лимфоидной ткани, наружная стенка которого образована ретикулярными клетками. Различают первичные и вторичные узелки. Первичные узелки – компактные гомогенные скопления В-лимфоцитов рециркулирующего пула, связанных с ретикулярными клетками и особым видом АПК. Имеются Т-клетки и макрофаги. Первичные узелки встречаются только во внутриутробном периоде. Под влиянием антигенов превращаются во вторичные. Вторичные узелки состоят из 1)короны и 2)герминативного центра Корона – скопление малых лимфоцитов на периферии узелка, полулунной формы на субкапсулярном полюсе. Содержит В-клетки рециркулирующего пула и В-клетки памяти. Герминативный центр – развивается только под влиянием антигенной стимуляции. Тут происходит АЗПД В-клеток в незрелые плазмоциты и В-клетки памяти. Из центра через глубокую кору в мозговые тяжи мигрируют незрелые плазмоциты. Межузелковая зона – содержит малые лимфоциты и макрофаги, замещается узелками при антигенной стимуляции. Глубокая кора (паракортикальная зона) – Т-зависимая зона. Дозревание Т-клеток из тимуса, а также их АЗПД. Образована диффузной лимфоидной тканью, представленной Т-клетками, лежащими в петлях ретикулярной ткани. Также есть АПК и плазмоциты. Лимфатические синусы – (промежуточные) и посткапиллярные венулы с высоким эндотелием, который способен взаимодействовать с хоминг-рецепторами Т- и В-лимфоцитов, обусловливая их миграцию из сосудистого русла. 18. Мозговое вещество лимфоузлов. Строение и клеточный состав. Система лимфатических синусов. Образовано ветвящимися и анастомозирующими тяжами лимфоидной ткани (мозговыми тяжами), между которыми располагаются трабекулы и мозговые лимфатические синусы. Мозговые тяжи – В-зависимая зона, содержат плазмоциты и В-лимфоциты с макрофагами. Лимфатические синусы – система особых внутриорганных лимфатических сосудов в корковом и мозговом веществе, обеспечивающая медленный ток лимфы через узел, в процессе которого она очищается на 99% от частиц и обогащается антителами, клетками лимфоидного ряда и макрофагами. (субкапсулярный -> промежуточный -> мозговой) 19. Селезенка. Строение и тканевой состав. В- и Т-зависимые зоны белой пульпы селезенки Селезенка –периферический и самый крупный орган иммунной системы. Формирует гуморальный и клеточный иммунитет, задерживает антигены, разрушает старые эритроциты и тромбоциты, депонирует крови и тромбоциты. Покрыта брюшиной и капсулой из ПСТ с ГМЦ. От капсулы отходят трабекулы. Паренхима (пульпа) включает 2 отдела с разными функциями: белую и красную пульпу. Белая пульпа (20%) – лимфоидная ткань, расположенная по ходу артерий (выполняет функцию: (а) улавливания из крови антигенов, (б) взаимодействие лимфоцитов с антигенами, АПК, (в) начальных этапов АЗПД), включает: (1) лимфатические узелки – располагаются по периферии ПАЛВ, сходны по структуре и функциям с лимфатическими узелками лимфатических узлов. Являются В-зависимой зоной селезенки. (2) ПАЛВ – окружают центральные артерии, состоят из цилиндрических компактных скоплений лимфоидной ткани, содержащей лимфоциты, макрофаги, ретикулярные и АПК. Являются Т-зависимой зоной селезенки. (3) маргинальную зону – располагается в виде тонкого слоя к периферии от ПАЛВ и узелков на границе белой и красной пульпы, рядом с маргинальным синусом и содержит лимфоциты, ретикулоциты и макрофаги. В наружной части накапливаются незрелые плазмоциты, мигрирующие в красную пульпу для дозревания. Служит местом нчального поступления в белую пульпу селезенки Т- и В-клеток и антигенов, которые здесь захватываются макрофагами. |