теория к биологии. био 9. 1. Плейотропное(множественное) действие генов
 Скачать 3.05 Mb. Скачать 3.05 Mb.
|

|
1.Плейотропное(множественное) действие генов.  Плейотропия – явление, при котором один ген обуславливает несколько признаков. Возможны случаи, когда при плейотропии ген по отношению к одному признаку доминирует, а по отношению к другому является рецессивным. В некоторых случаях при плейотропии наблюдается расщепление при скрещивании моногибридов не 3 : 1, а 2 : 1. Это объясняется тем, что один из признаков определяется летальным геном, т. е. в гомозиготе ведет к гибели. Плейотропия – явление, при котором один ген обуславливает несколько признаков. Возможны случаи, когда при плейотропии ген по отношению к одному признаку доминирует, а по отношению к другому является рецессивным. В некоторых случаях при плейотропии наблюдается расщепление при скрещивании моногибридов не 3 : 1, а 2 : 1. Это объясняется тем, что один из признаков определяется летальным геном, т. е. в гомозиготе ведет к гибели.  Примеры: у многих растений ген, обуславливающий окраску цветка, вызывает изменение окраски других органов (стебля, листьев, семян).- Рис. 1. Львиный зев у человека мутантный ген, контролирующий развитие соединительной ткани, обуславливает синдром «паучьи пальцы» (синдром Марфана) и вызывает не только удлинение пальцев рук и ног, но также изменение формы лица, дефект хрусталика, врождённые пороки сердца и др. 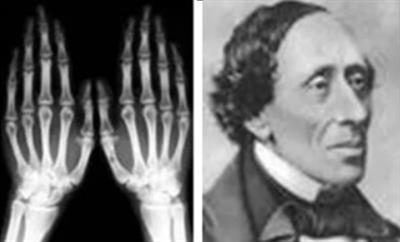 ген, отвечающий за рыжий цвет волос, обуславливает также более светлую окраску кожи и появление веснушек. 2. Количественная и качественная специфика проявления гена в признак: пенетрантность и экспрессивность. Экспрессивность - качественный показатель фенотипического проявления гена, характеризует степень выраженности признака. Степень проявления фенотипа – экспрессивность. Образно ее можно сравнить со степенью тяжести болезни в клинической практике. Экспрессивность подчиняется законам распределения Гаусса (некоторые в малом или среднем количестве). В основе изменчивости экспрессивности лежат и генетические факторы, и факторы внешней среды. Экспрессивность – очень важный показатель фенотипического проявления гена. Количественно ее степень определяют, используя статистический показатель. При отсутствии изменчивости признака, контролируемого данным аллелем, говорят о постоянной экспрессивности, в противном случае – об изменчивой или вариабельной. Группа крови – признак с постоянной экспрессивностю, цвет глаз – признак с вариабельной экспрессивностью. Экспрессивность также является показателем, характеризующим фенотипическое проявление наследственной информации. Она характеризует степень выраженности признака и, с одной стороны, зависит от дозы соответствующего аллеля гена при моногенном наследовании или от суммарной дозы доминантных аллелей генов при полигенном наследовании, а с другой — от факторов среды. Примером служит интенсивность красной окраски цветков ночной красавицы, убывающая в ряду генотипов АА, Аа, аа, или интенсивность пигментации кожи у человека, увеличивающаяся при возрастании числа доминантных аллелей в системе полигенов от 0 до 8 (см. рис. 3.80). Влияние средовых факторов на экспрессивность признака демонстрируется усилением степени пигментации кожи у человека при ультрафиолетовом облучении, когда появляется загар, или увеличением густоты шерсти у некоторых животных в зависимости от изменения температурного режима в разные сезоны года. Генетический признак может даже не проявляться в некоторых случаях. Если ген есть в генотипе, но он вовсе не проявляется – он пенетрирован. (русский ученый Тимофеев-Рисовский 1927 год). Пенетрантность – количество особей (%), проявляющих в фенотипе данный ген, по отношению к количеству особей, у которых этот признак мог бы проявиться. Пенетрантность свойственна проявлению многих генов. Важен принцип – «все или ничего» - либо проявляется, любо нет. Пенетрантность- способность гена проявить свое действие в фенотипе. Выражается в виде процентного отношения количества организмов, у которых данный ген фенотипически проявляется к общему числу организмов, имеющих этот ген. Если ген проявляется в фенотипе у всех его носителей, то пенетрантность гена равна 100% (полная пенетрантность). - наследственный панкреатит – 80% - вывих бедра – 25% - пороки развития глаз - ретинобластома – 80% - отосклероз – 40% - колотокома – 10% - Пенетрантностъ отражает частоту фенотипического проявления имеющейся в генотипе информации. Она соответствует проценту особей, у которых доминантный аллель гена проявился в признак, по отношению ко всем носителям этого аллеля. Неполная пенетрантность доминантного аллеля гена может быть обусловлена системой генотипа, в которой функционирует данный аллель и которая является своеобразной средой для него. Взаимодействие неаллельных генов в процессе формирования признака может привести при определенном сочетании их аллелей к непроявлению доминантного аллеля одного из них. Хорея Гентингтона проявляется в непроизвольном подергивании головы. Конечностей, постепенно прогрессирует и приводит к смерти. Может проявиться в раннем постэмбриональном периоде, в зрелом возрасте или не проявиться вообще. И экспрессивность, и пенетрантность поддерживаются естественным отбором, т.е. гены, контролирующие патологические признаки могут иметь разную экспрессивность и пенетрантность: заболевают не все носители гена, а у заболевших степень проявления будет различна. Проявление или неполное проявление признака, а так же его отсутствие зависит от среды и от модифицирующего действия других генов. Ген может действовать плейотропно (множественно), т.е. опосредовано влиять на течение разных реакций и развитие многих признаков. Гены могут оказывать влияние на другие признаки на разных стадиях онтогенеза. Если ген включается в позднем онтогенезе, то оказывается незначительное действие. Если на ранних стадиях – изменения более значительны. 3.Роль наследственных факторов и среды в развитии заболеваний Фенотип человека, формирующийся на различных стадиях его онтогенеза, так же как фенотип любого живого организма, является в первую очередь продуктом реализации наследственной программы. Степень зависимости результатов этого процесса от условий, в которых он протекает, у человека определяется его социальной природой. Наследственные болезни. Развитие этих заболеваний целиком обусловлено дефектностью наследственной программы, а роль среды заключается лишь в модифицировании фенотипических проявлений болезни. К этой группе патологических состояний относят хромосомные болезни, в основе которых лежат хромосомные и геномные мутации, и моногенно наследуемые заболевания, обусловленные генными мутациями. В качестве примера можно назвать болезнь Дауна, гемофилию, фенилкетонурию. Наследственные болезни всегда связаны с мутацией, однако фенотипическое проявление последней, степень выраженности патологических симптомов у разных индивидумов могут различаться. Мультифакториальные заболевания, или болезни с наследственным предрасположением. К ним относится большая группа распространенных заболеваний, особенно болезни зрелого и преклонного возраста, такие, как гипертоническая болезнь, ишемическая болезнь сердца, язвенная болезнь желудка и двенадцатиперстной кишки и т.д. Причинными факторами их развития выступают неблагоприятные воздействия среды, однако реализация этих воздействий зависит от генетической конституции, определяющей предрасположенность организма. Соотносительная роль наследственности и среды в развитии разных болезней с наследственным предрасположением неодинакова. +Лишь немногие формы патологии обусловлены исключительно воздействием факторов среды— травма, ожог, обморожение, особо опасные инфекции. Но и при этих формах патологии течение и исход заболевания в значительной степени определяются генетическими факторами. Болезнь, как и жизнедеятельности организма, являются результатом взаимодействия наследственных и средовых факторов. Если внутренние факторы наследственно изменены, то возникает патологический процесс. Факторы внутренней среды - это в конечном счете результат взаимодействия генетических и средовых факторов в онтогенезе, потому что уровень гормонов в организме, особенности обмена веществ, иммунные реакции исходно определяются функционированием соответствующих генов, другими словами, генетической конституцией. Наследственные факторы, определяющие основу внутренней среды организма в широком смысле слова, принимают самое непосредственное участие в формировании патологических процессов, либо выступая в роли этиологического фактора, либо участвуя в патогенезе заболевания. С генетической точки зрения все болезни в зависимости от относительной значимости наследственных и средовых факторов в их развитии можно разделить на наследственные болезни, болезни с наследственной предрасположенностью и ненаследственные болезни (рис. 2.3). Наследственные болезни вызваны мутациями. Проявление патологического действия мутации как этиологического фактора практически не зависит от среды. Среда может только менять выраженность симптомов болезни и тяжесть ее течения. К заболеваниям этой группы относятся хромосомные и генные наследственные болезни. (болезнь Дауна, нейрофиброматоз, гемофилия, фенилкетонурия, муковисцидоз, ахондроплазия и др.). О болезнях с наследственной предрасположенностью говорят тогда, когда болезнь развивается у лиц с определенной генетической характеристикой под влиянием факторов окружающей среды. Эти болезни называют также мультифакториальными. Наследственность служит и этиологическим, и патогенетическим фактором. Для пенетрантности мутантных генов необходим соответствующий фактор окружающей среды. К таким заболеваниям относятся, например, некоторые формы подагры, диабета, фармако- и экогенетические болезни. Подобные заболевания развиваются после контактов с проявляющим болезнь внешним фактором, специфичным для каждого мутантного гена. (атеросклероз, гипертоническая болезнь, туберкулез, экзема, псориаз, язвенная болезнь) В происхождении ненаследственных болезней определяющую роль играет среда. Сюда относится большинство травм, инфекционных болезней, ожогов и т.д. Генетические факторы могут влиять только на течение патологических процессов (выздоровление, восстановительные процессы, компенсация нарушенных функций). 4. Строение и типы хромосом. Понятие о гетеро- и эухроматине. Х 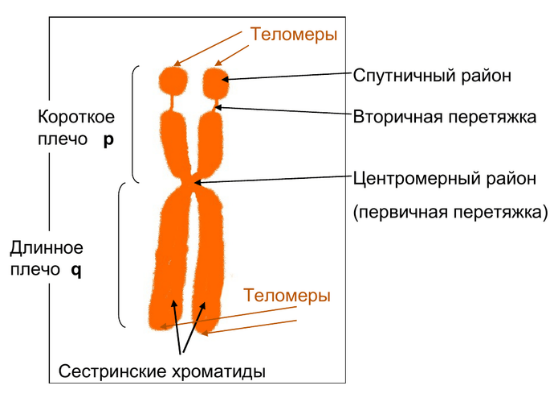 ромосомы— нуклеопротеидные структуры в ядре эукариотической клетки (клетки, содержащей ядро), которые становятся легко заметными в определённых фазах клеточного цикла (во время митоза или мейоза). Хромосомы представляют собой высокую степень конденсации хроматина, постоянно присутствующего в клеточном ядре. Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия всё чаще говорят о бактериальных хромосомах. В хромосомах сосредоточена большая часть наследственной информации. ромосомы— нуклеопротеидные структуры в ядре эукариотической клетки (клетки, содержащей ядро), которые становятся легко заметными в определённых фазах клеточного цикла (во время митоза или мейоза). Хромосомы представляют собой высокую степень конденсации хроматина, постоянно присутствующего в клеточном ядре. Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия всё чаще говорят о бактериальных хромосомах. В хромосомах сосредоточена большая часть наследственной информации. Первичная перетяжка Хромосомная перетяжка, в которой локализуется центромера и которая делит хромосому на плечи. Вторичные перетяжки Морфологический признак, позволяющий идентифицировать отдельные хромосомы в наборе. От первичной перетяжки отличаются отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают корот кими и длинными и локализуются в разных точках по длине хромосомы. У человека это 13, 14, 15, 21 и 22 хромосомы. Теломеры – концевые участки хромосом, содержащие до 10 тысяч пар нуклеотидов с повторяющейся последовательностью ТТАГГГ. Теломеры не содержат генов, они защищают концы хромосом он действия нуклеаз – ферментов, разрушающих ДНК обеспечивают прикрепление концов хромосом изнутри к ядерной оболочке защищают гены от концевой недорепликации. Спутник– это хромосомный сегмент, чаще всего гетерохроматический, расположенный дистально от вторичной перетяжки. По классическим определениям спутник – сферическое тельце с диаметром, равным диаметру хромосомы или меньше его, которое связано с хромосомой тонкой нитью. Выделяют следующие 5 типов спутников: микроспутники – сфероидальной формы, маленькие спутники с диаметром вдвое или еще меньше диаметра хромосомы; макроспутники – довольно крупные формы спутников с диаметром, превышающим половину диаметра хромосомы; линейные С. - спутники, имеющие форму длинного хромосомного сегмента. Вторичная перетяжка значительно удалена от терминального конца; терминальные С. – спутники, локализованные на конце хромосомы; интеркалярные С. – спутники, локализованные между двумя вторичными перетяжками. Типы строения хромосом 1.телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце); 2.акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом); 3.субметацентрические (с плечами неравной длины, напоминающие по форме букву L); 4.метацентрические (V-образные хромосомы, обладающие плечами равной длины). Хроматин - основной компонент клеточного ядра. В среднем в хроматине 40% приходится на ДНК и около 60% на белки. В структурном отношении хроматин представляет собой нитчатые комплексные молекулы дезоксирибонуклеопротеида, которые состоят из ДНК, ассоциированной с гистонами и иногда еще с негистоновыми белками. Способность к дифференциальному окрашиванию легла в основу выявления двух фракций хроматина – гетеро – и эухроматина. Хейтц, открывший это явление, нашел, что определенные участки хромосом остаются в конденсированном состоянии в течении всего клеточного цикла и назвал их гетерохроматин, а участки, деконденсирующиеся в конце митоза и слабо окрашенные – эухроматином. Гетерохроматиновые участки функционально менее активны, чем эухроматиновые, в которых и локализована большая часть известных генов. Однако, гетерохроматин имеет определенное генетическое влияние; к примеру, определяющие пол хромосомы не могут рассматриваться как генетически неактивные, хотя они часто полностью состоят из гетерохрома тина. Кроме того, установлено, что стабильность генетического выражения эухроматина обуславливается близостью к гетерохроматину. Цитогенетические методы исследования хромосом Цитогенетический метод включает два основных вида исследования: 1) изучение хромосомного набора в соматических клетках организма человека, т.е. кариотипа; 2) определение полового хроматина. 1. Исследование кариотипа. Бурное развитие этот метод получил после 1956 года, когда шведские ученые Дж. Тийо и А. Леван предложили новую методику исследования хромосомного набора и установили, что кариотип человека в норме содержит 46 хромосом. Для исследования берут 1 мл крови, выделяют из нее лимфоциты и культивируют их на питательной среде. Через определенное время воздействуют на культуру клеток колхицином, который останавливает деление лимфоцитов на стадии метафазы. Клеточную суспензию наносят на предметные стекла, окрашивают и микроскопируют. Изучению подвергаются метафазные пластинки. Микропрепарат фотографируют, делают отпечатки на фотобумаге, вырезают изображение каждой хромосомы ножницами и наклеивают на белую бумагу в ряд попарно, начиная с первой пары гомологов и заканчивая парой половых хромосом. Такое расположение хромосомного набора называетсяидиограммой. С конца 60-х годов стали применяться методы дифференциального окрашивания хромосомных препаратов, которые позволяют точно идентифицировать каждую хромосому в наборе и диагностировать структурные аберрации. Метод позволяет поставить диагноз хромосомного заболевания человека. 2. Исследование полового хроматина (телец Барра) – экспресс-метод. При помощи шпателя делают соскоб со слизистой щеки, наносят мазок на предметное стекло, окрашивают и исследуют под микроскопом клетки, находящиеся на стадии интерфазы. Метод позволяет установить количество Х-хромосом в кариотипе по числу телец Барра в клетке. В норме у женщин одна из Х-хромосом в период интерфазы не функционирует и образует тельце Барра (половой хроматин), которое хорошо видно в микроскоп как глыбка хроматина, прилежащая к ядерной мембране. В мужских соматических клетках тельце Барра в норме отсутствует. Понятие о кариотипе и идиограмме. Денверская и Парижская классификация хромосом. Хромосомный набор (кариотип) соматической клетки характеризуется формой хромосом, их количеством, размерами, характерными для каждого вида. Денверская классификация хромосом, которая помимо размеров хромосом, учитывает их форму, положение центромеры и наличие вторичных перетяжек и спутников. 23 пары хромосом человека разбили на 7 групп от A до G. Важным параметром является центромерный индекс (ЦИ), который отражает отношение (в %) длины короткого плеча к длине всей хромосомы. К группе A относят 1-3 хромосомы. Это большие метацентрические и субметацентрические хромосомы, их центромерный индекс от 38-49. Группа B (4 и 5 пары). Это большие субметацентрические хромосомы, ЦИ 24-30. Группа C (6-12 пары). Хромосомы среднего размера, субметацентрические, ЦИ 27-35. К этой группе относят и Х-хромосому. Группа D (13-15 пары). Хромосомы акроцентрические, сильно отличаются от всех других хромосом человека, ЦИ около 15. Группа E (16-18 пары). Относительно короткие, метацентрические или субметацентрические, ЦИ 26-40. Группа F (19-20 пары): две короткие, субметацентрические хромосомы, ЦИ 36-46. Группа G (21-22 пары): это маленькие акроцентрические хромосомы, ЦИ 13-33. К этой группе относят и Y-хромосому. Парижская классификация хромосом В начале 70-х годов XX века был разработан метод дифференциальной окраски хромосом, выявляющий характерную сегментацию, который позволил индивидуализировать каждую хромосому. Каждая хромосома человека содержит свойственную только ей последовательность полос, что позволяет идентифицировать каждую хромосому. Хромосомы спирализованы максимально в метафазе, менее спирализованы в профазе и прометафазе, что позволяет выделить большее число сегментов, чем в метафазе.На метафазной хромосоме приводятся символы, которыми принято обозначать короткое и длинное плечо, а также расположение районов и сегментов. При описании кариотипа: • указывается общее число хромосом и набор половых хромосом, между ними ставится запятая (46, XX; 46, XY); • отмечается какая хромосома лишняя или какой не хватает (это указывается ее номером 5, 6 и др., или буквами данной группы А, В и др.); знаком «+» указывают на увеличение количества хромосом, знаком «-» указывают на отсутствие данной хромосомы 47, XY,+ 21; • плечо хромосомы, в котором произошло изменение (удлинение короткого плеча указывается символом (р+); укорочение (р-); удлинение длинного плеча указывается символом (q+); укорочение (q-); • символы перестроек (транслокация обозначается t, а делеция — del) помещают перед номерами вовлеченных хромосом, а перестроечные хромосомы заключают в скобки. Наличие двух структурно-аномальных хромосом обозначается точкой с запятой (;) или нормальной дробью (15/21). Система записи кариотипов 46, XX — нормальный кариотип (женщина) 46, XY — нормальный кариотип (мужчина) 45, X — синдром Шерешевского-Тернера 47 XXY 1 46, xxxY I — синдром Клайнфельтера 47, XXX — синдром «трисомии по Х-хромосоме» 47, XYY — синдром Вай-Вай 47, XX, + 21 — синдром Дауна (женщина) 47, XY, + 21 — синдром Дауна (мужчина) 47, XX, + 18 — синдром Эдвардса (женщина) 47, XY, + 18 — синдром Эдвардса (мужчина) 47, XX, + 13 — синдром Патау (женщина) 47, XY, + 13 — синдром Патау (мужчина) 46, XX, del (5p-) — синдром кошачьего крика (женщина) 46, XY, del (5р-) — синдром кошачьего крика (мужчина) 46, XX, del (13q-) — синдром Орбели (женщина) 46, XY, del (13q-) — синдром Орбели (мужчина) Сегменты и районы метафазной хромосомы обозначаются цифрами, центромера служит исходной точкой для цифровой схемы. При определении локализации гена используют 4 критерия: номер хромосомы, символ плеча, номер района и номер сегмента в пределах этого района. Например, запись 1р32 означает, что речь идет о хромосоме первой пары, коротком плече, районе 3, сегменте 2. Для гена Rh формула локализации: 1р35. Принципы составления идиограмм +Если расположить фотографии гомологичных хромосом по мере возрастания их размеров, то можно получить так называемую идиограмму кариотипа. Таким образом, идиограмма - это графическое изображение хромосом. На идиограмме пары гомологов располагаются рядами в порядке убывающего размера. 5. Цитогенетический метод (кариотипирование). Цитогенетический метод С помощью данного метода можно изучать наследственный материал клетки: совокупность хромосом в целом (кариотипирование) или наличие и количество Х-хромосом (определение полового хроматина — число глыбок полового хроматина или телец Барра). Исследование проводится с помощью светового микроскопа (изготовление и изучение микропрепаратов). Кариотипирование На рис. представлена последовательность действий для получения кариотипа человека. 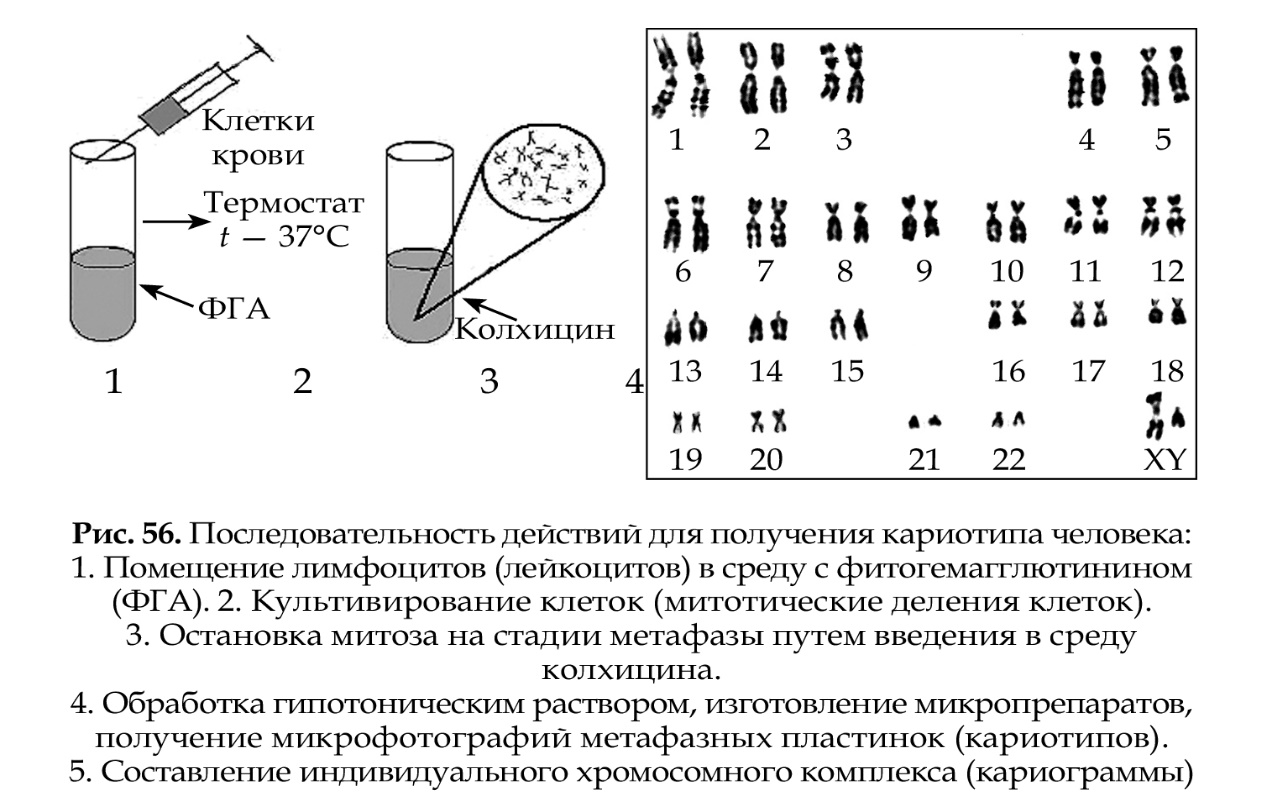   Понятие о кариотипе и идиограмме. Денверская и Парижская классификация хромосом. Кариотип – это совокупность хромосом диплоидной клетки данного биологического вида. Характеристика кариотипа:число,размер,форма. 46 хромосом:22 пары аутосомы и 1 пара гоносом. Исследуемая фаза:метафаза исследуемые клетки:лимфоциты периферической крови. Этапы кариотипирования: Цитогенетический метод. 1)лимфоциты+фитогемаглютинин->увеличколичесвоклеток,вступивших в митоз; 2).добавляем колхицин->разрушение веретена деления->остановка клеток в метафазе; 3).+гипотонический раствор->хромосомы отделяются друг от друга 4)фиксация этанол-уксусным раствором 5).окрашивание хромосом: а).краситель романовского-Гимзы-хромосомы окрашивравнгомерно б).краситель Гимзы и др. Хромосомы окрашив дифференцированно. Спектральное кариотипирование-окрашивание флюурисцентнымикрасителями,специф для каждой пары гомологичных хромосом.Метод обличает выявление пар хромосом и межхромосомныхтранслокаций. Изучение прометафазныххромосом.исслед фаза-прометафаза,хромос более расплетенные->выявление более мелких хромосомных нарушений. Использование Днк-зондов(FISH).исслед фаза-интерфаза или метафаза. Идиограмма-это систематизированный кариотип,в котором хромосомы рапсолагаются по мере убывания величины. Норм идиограммачеловека,согласноденферской и парижской классифвключ 7 гр хромосом(от А доG) Денверская(1960): рутинная(равномерная) окраска хромосом размеры и формы хромосом,положениецентромер,наличие вторичных перетяжек и спутников. Метод позволяет выявить числовые мутации и крупные хромосом мутации,установитьпол,однако не позволяет дифференциировать хромосомы внутри 1 гурппы. Парижская (1971) Дифференциированная окраска Поперечная исчерченность-чередование гетеро-хроматиновых районов каждой хромосомы одинаковы у гомологич хромосом->точная идентификация хромосом. Позволяет выявить численные нарушения,пол,а также большинство мелких и крупных хром мутаций. Группа А-1-3 пары.1 и 3-крупные метацентрические,2-крупные субметацентрические 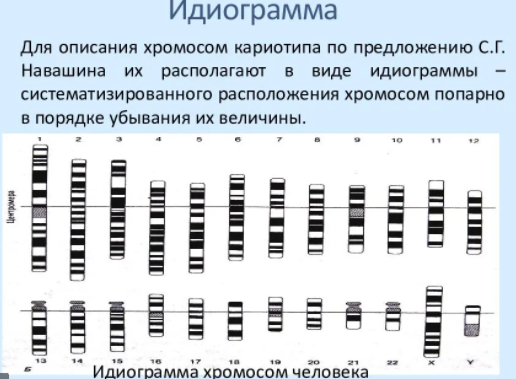 Группа В-4 и 5.крупныесубметацентрические Группа В-4 и 5.крупныесубметацентрическиегруппа С – 6-12,Х. Средние субметацентрические Группа D- 13-15.Средниеакроцентрические Группа Е-16-18.Мелкиесубметацентрические Группа F – 19,20. Мелкие метацентрические. +Группа G-21,22 и У. Мелкиеакрочентрические.   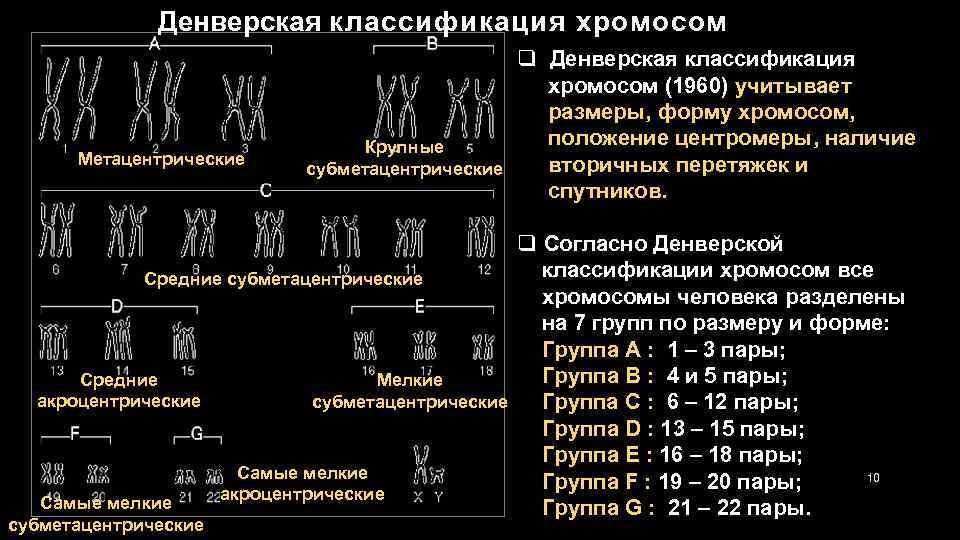  7.Кариотип человека в норме и при патологии. Примеры +Хромосомные болезни возникают при нарушении кариотипа (нормальный кариотип человека 46,ХХ, 46,ХУ), которое может быть вызвано изменением числа (геномные мутации) или структуры (хромосомные мутации) аутосом и половых хромосом. При хромосомных болезнях имеется определенный комплекс стабильных аномальных признаков - симптомов, который входит в понятие синдром. Синдромы характеризуются определенной частотой проявления, продолжительностью жизни детей и средним весом при рождении, внешними морфологическими признаками, пороками развития внутренних органов, функциональными симптомами, дерматоглификой и определенным кариотипом. Среди хромосомных болезней, связанных с изменением числа половых хромосом (моносомии, полисомии), наиболее часто встречаются синдромы Клайнфельтера (47,ХУ), Шерешевского-Тернера (45,Х), трисомия по Х-хромосоме (47,ХХХ), полисомия по У-хромосоме (47,ХУУ); может наблюдаться полисомия по Х-хромосоме и У-хромосоме одновременно ( 48,ХХУУ). Эффект мутаций аутосом различен: при геномных мутациях 1-12 хромосом возникают аномалии, несовместимые с жизнью; 13-18 - полулетальные мутации (спонтанные аборты, множественные уродства, незначительная продолжительность жизни родившихся от нескольких недель до нескольких лет - синдром Патау (47,ХХ/ХУ + 13), синдром Эдвардса (47,ХУ/ХХ + 18). Трисомия по 21 хромосоме - синдром Дауна (47,ХХ/ХУ + 21) - является аномалией, совместимой с жизнь. При анализе синдромов по половым хромосомам и аутосомам показано, что при аномалиях по половым хромосомам сохраняется нормальный интеллект или отмечается его снижение, но в большей степени нарушается развитие половых органов и гормонозависимый рост (выше или ниже средней нормы). Моносомия Х встречается реже (1: 25ОО), чем полисомии ХХУ (1: 700) и ХХХ (1: 1000). Хромосомные аберрации в основном представлены делециями и транслокациями: при делеции короткого плеча 5 хромосомы (46,ХХ/ХУ 5р-) наблюдается синдром «кошачьего крика» (название обусловлено сходством плача ребенка с мяуканьем кошки), происхождение которого вызвано нарушением центральной нервной системы, а не аномалией голосового аппарата. Встречаются делеции по 13, 18, 21, 22 хромосомам (46,ХХ/ХУ 13q- - синдром Орбели). Транслокация 15/21 хромосом приводит к возникновению синдрома Дауна; а 9/22- хроническому миелолейкозу. Для диагностики хромосомных болезней используется цитогенетический метод (кариотипирование, определение полового хроматина); методы амниоцентеза и дерматоглифики. Хромосомные болезни не наследуются, так как у больных нарушена репродуктивная функция, но синдромы появляются в каждом поколении с определенной частотой как результат вновь возникших мутаций у здоровых людей. 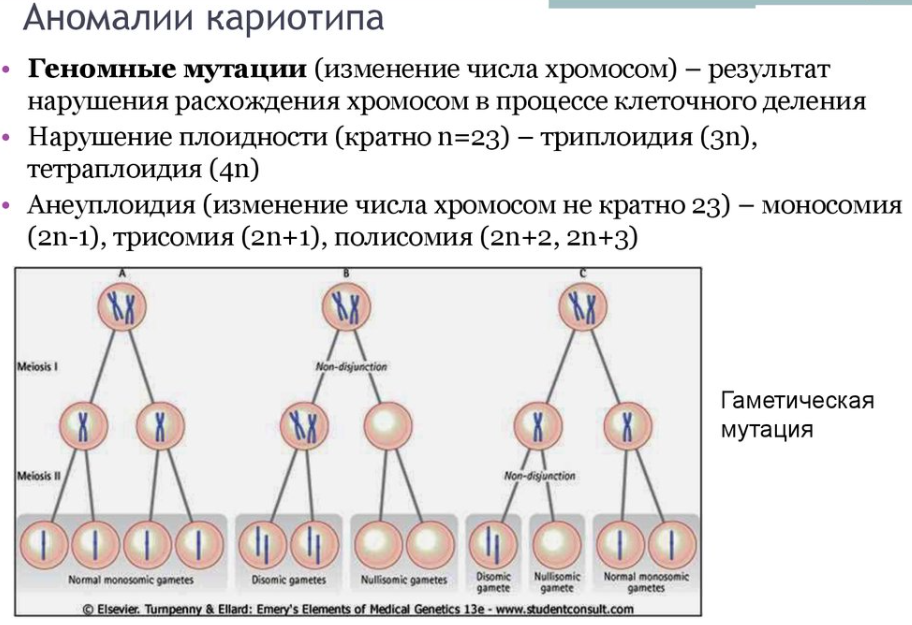 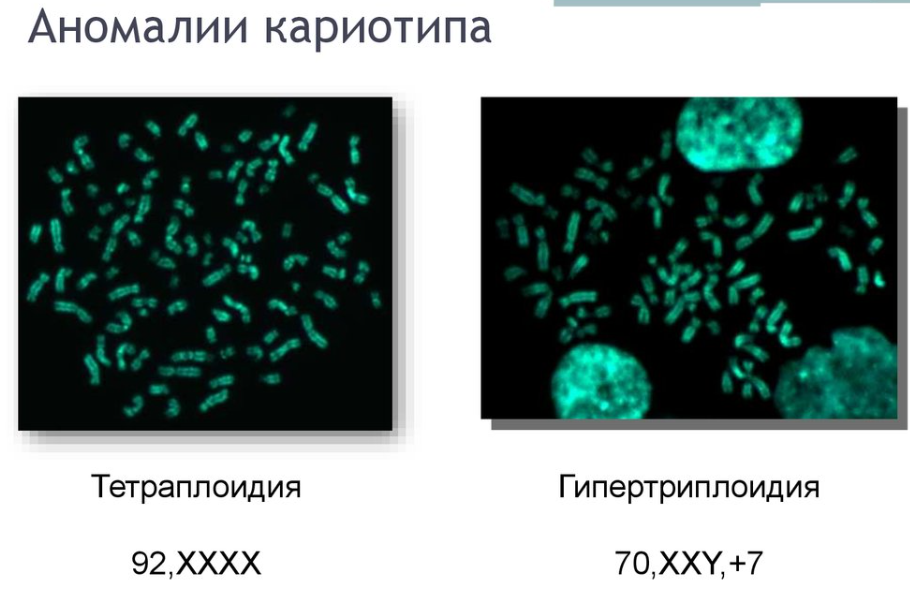  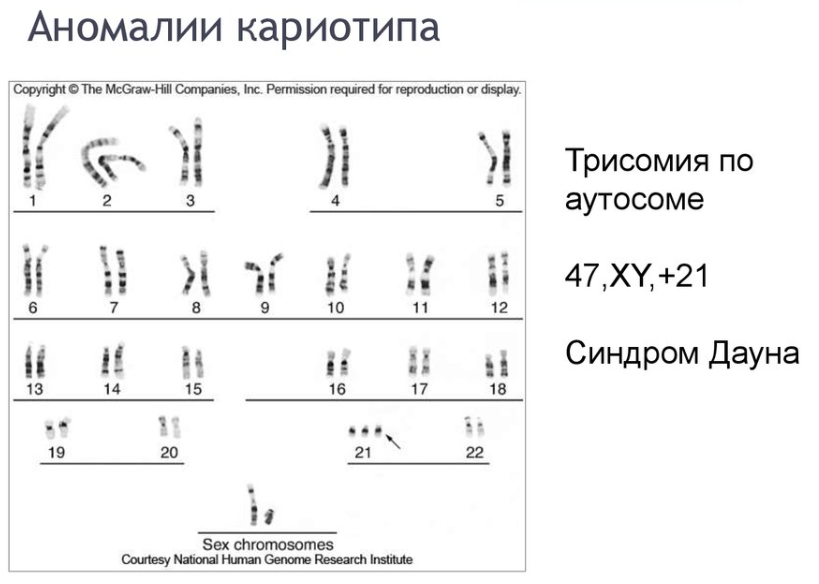 8.Работы Моргана по изучению полного и неполного сцепления генов.Основоположник хромосомной теории Томас Гент Морган, американский генетик, нобелевский лауреат, выдвинул гипотезу об ограничении законов Менделя. В экспериментах он использовал плодовую мушку-дрозофилу, обладающую важными для генетических экспериментов качествами: неприхотливостью, плодовитостью, небольшим количеством хромосом (четыре пары), множеством четко выраженных альтернативных признаков. +Работы Моргана заложили основы хромосомной теории наследственности, они показали, что ограничения в свободной комбинаторике некоторых генов обусловлены расположением этих генов в одной хромосоме и их физическим сцеплением. Морганом было установлено, что сцепление генов, расположенных в одной хромосоме, не является абсолютным. Во время мейоза хромосомы одной пары могут обмениваться гомологичными участками между собой с помощью процесса, который называется кроссинговером. Чем дальше друг от друга расположены гены в хромосоме, тем чаще они разделяются кроссинговером. На основе этого феномена была предложена мера силы сцепления генов — процент кроссинговера — и построены первые генетические карты хромосом для разных видов дрозофилы. В качестве объекта генетического анализа была выбрана плодовая мушка дрозофила и Морган изучал наследование у нее разных признаков. Скрестив гомозиготную самку с серыми телом и длинными крыльями (домин), с гомозиготным чернокрылым короткокрылым самцом, в F1 – однообразие (серое тело, длинные крылья) Далее Морган провел несколько анализирующих скрещиваний. Оказалось, что результаты будут разные в зависимости от пола гибрида. Если гибридным был самец, то в потомстве получалось 2 фенотипических класса полностью повторяющих признаки родителей. Если гибридной была самка, то получалось 4 фенотипических классов потомком в неравных пропорциях. Большую часть потомства (83%) составляют потомки с родительскими признаками, меньшую (17%) – особи с новыми комбинациями признаков. Морган сделал вывод, что сцепление может быть неполным, где группа сцепления нарушается кроссинговером. Необычность процентного соотношения у потомков объясняется тем, что кроссинговер происходит не всегда, частота кроссинговера зависит от расстояния между генами – чем больше расстояние, тем меньше силы сцепления между генами, тем чаще кроссинговер. Гаметы, в которые попали хромосомы, не прошедшие кроссинговер, называются некроссоверные. Если в гаметах хромосомы претерпевшие кроссинговер – кроссоверные. Морган и его ученики установили следующее: 1. Гены, расположенные в одной хромосоме, наследуются совместно или сцепленно. 2. Группы генов, расположенных в одной хромосоме, образуют группы сцепления. Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и п+1 у гетерогаметных особей. 3. Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссинговера возникают гаметы, хромосомы которых содержат новые комбинации генов. 4. Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера. За единицу расстояния между генами принимают 1 морганиду (1% кроссинговера) или процент появления кроссоверных особей. При значении этой величины в 10 морганид можно утверждать, что частота перекреста хромосом в точках расположения данных генов равна 10% и что в 10% потомства будут выявлены новые генетические комбинации. 5. Для выяснения характера расположения генов в хромосомах и определения частоты кроссинговера между ними строят генетические карты. Карта отражает порядок расположения генов в хромосоме и расстояние между генами одной хромосомы. Эти выводы Моргана и его сотрудников получили название хромосомной теории наследственности. Важнейшими следствиями этой теории являются современные представления о гене как о функциональной единице наследственности, его делимости и способности к взаимодействию с другими генами. Опыты Томаса Моргана (дигибридное скрещивание) 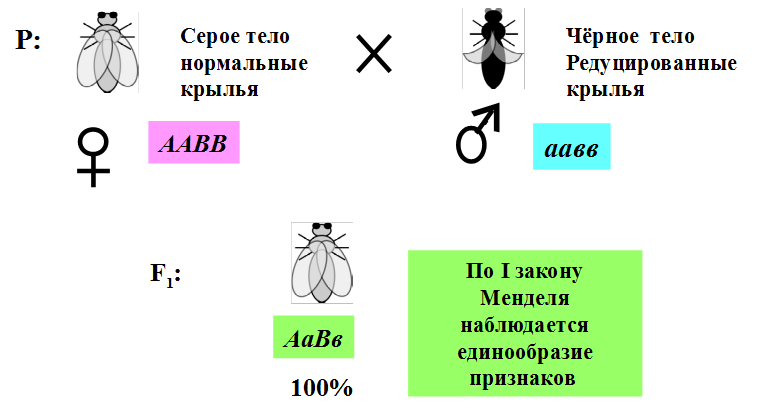 Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых).При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1.  Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями).  Давайте разбираться. Цитологические основы закона сцепленного наследования признаков Томаса Моргана. 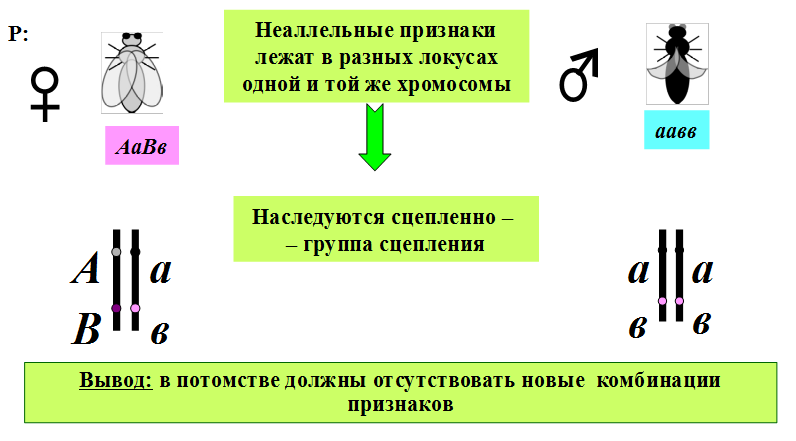 Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип АаВb и ааbb 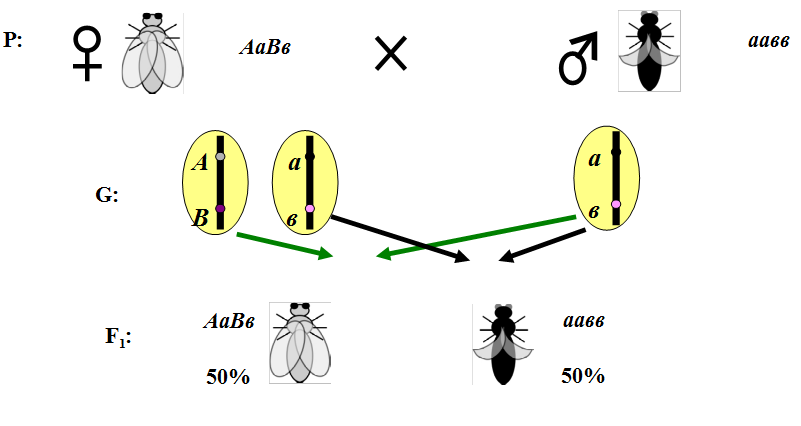 Так откуда же появились новые комбинации признаков в потомстве? Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1. 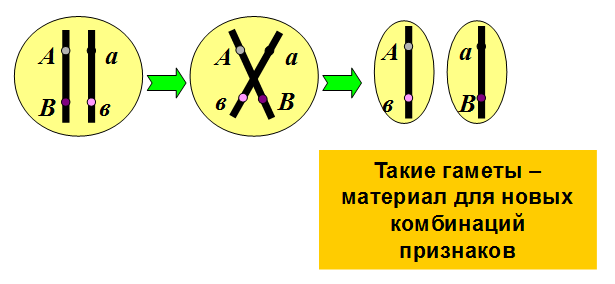 Ещё раз вспомним, что такое КРОССИНГОВЕР (англ. crossing-over — перекрест или пересечение) — процесс обмена гомологичных хромосом участками во время их конъюгации в профазе I мейоза. Кроссинговер является одним из механизмов генетической рекомбинации (обмена генами). Частота его зависит от расстояния между генами: чем дальше расположены гены друг от друга, тем чаще между ними идет перекрест. 1% кроссинговера принят за единицу расстояния между генами. Она названа морганидой в честь Т. Моргана, разработавшего принципы генетического картирования. Морганида — условная единица расстояния между генами в хромосомах. Соответствует расстоянию, при котором кроссинговер происходит в 1% гамет, т. е. 1 морганида эквивалентна 1% кроссоверных гамет. Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом. Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними. Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом. Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.  Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.  Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей. Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей. Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза). 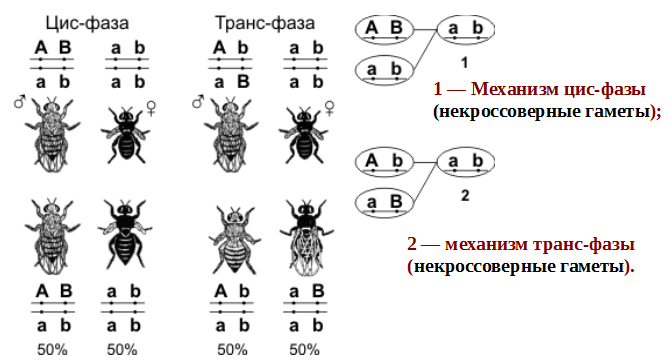 Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности: - гены располагаются в хромосомах; - различные хромосомы содержат неодинаковое число генов; - набор генов каждой из негомологичных хромосом уникален; - каждый ген имеет определенное место (локус) в хромосоме; - в идентичных локусах гомологичных хромосом находятся аллельные гены; - гены расположены в хромосомах в определенной линейной последовательности; - гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов; - сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; - частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера; - каждый вид имеет характерный только для него набор хромосом — кариотип. Генетические карты хромосом Генетическая карта хромосомы схематическое изображение хромосом с указанием мест фактического размещения отдельных генов. Для соcтавления генетических карт хромосом необходимо выявление множество мутантных генов и проведения многочисленных скрещиваний. Расстояние между генами на генетической карте хромосом определяют по чистоте кроссинговера между ними. Единицей расстояния генетической карте хромосом мейотически делящихся клеток является морганида, соотвеьсвующая одному проценту кроссинговера. В норме генетические карты хромосом у эукариот линейные. 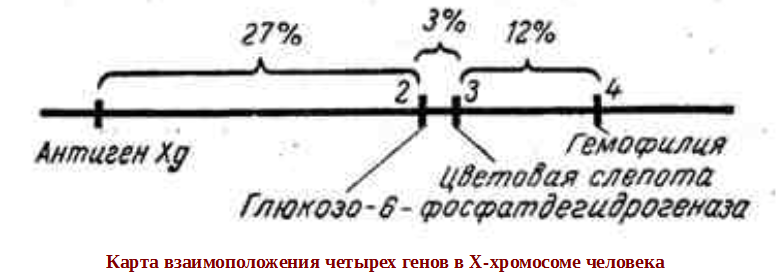 У прокариот и вирусов генетические карты хромосом также строят с помощью рекомбинации. При картировании генов у бактерий с помощью конъюгации получается кольцевая генетическая карта хромосомы. З 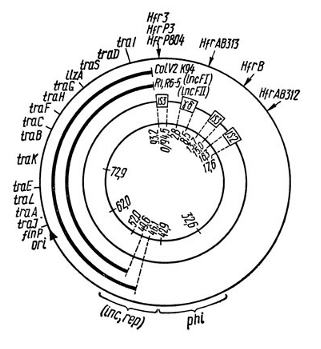 начение генетических карт позволяет планировать работу по получению организмов с определенными сочетаниями признаков, что используется в генетических экспериментах селекционной практике. На основе же генетических карт проводят генетический анализ. начение генетических карт позволяет планировать работу по получению организмов с определенными сочетаниями признаков, что используется в генетических экспериментах селекционной практике. На основе же генетических карт проводят генетический анализ.Ц  итологическая карта хромосомы представляет собой фотографию или точный рисунок хромосомы, на котором отмечается последовательность расположения генов. Ее строят на основе сопоставления результатов анализирующего скрещивания и хромосомных перестроек. итологическая карта хромосомы представляет собой фотографию или точный рисунок хромосомы, на котором отмечается последовательность расположения генов. Ее строят на основе сопоставления результатов анализирующего скрещивания и хромосомных перестроек.8.Основные положения хромосомной теории За ее развитие и доказательство великому генетику Томасу Моргану в 1933 г. была присвоена Нобелевская премия. 1. Хромосомы являются носителями наследственной информации. 2. Каждая хромосома содержит множество генов. 3. Гены, расположенные в одной хромосоме, наследуются совместно (сцепленно) и образуют группу сцепления. 4.Число групп сцепления равно гаплоидному числу хромосом данного вида (у гетерогаметного пола больше на одну). 5. Сцепление аллелей, расположенных в одной хромосоме, может нарушаться за счет кроссинговера. 6. Гены расположены в хромосомах в линейной последовательности. 7. Частота кроссинговера (нарушения сцепления) пропорциональна расстоянию между генами. 9. Понятие о картировании хромосом. Методы картирования хромосом человека, генетические карты (физические, химические) Картирование – это получение графического изображения хромосомы с точной локализац генов на данной графи карте. Генетическая карта-это отрезок прямой,на которой указываютмягены,расст между ними в морганидах или процентах кроссинговера. Основана на гибридологическом методе,поэтому к человеку не применима. Анализ родословных Гибридизация соматических клеток Цитологическая карта основана накариотипировании и представляет собой точную фотографию или рис. Хромосомы. +Физическая карта-наиболее точная карта,гдеиспольз гены с расст между ними в парах. Химическая карта—полинуклеотиды расположены от одной теломеры до другой    |