Раздел 1. Микробиология. 1. Предмет и з
 Скачать 7.68 Mb. Скачать 7.68 Mb.
|

|
В состав белков входят как обычные для эукариотов аминокислоты, так и оригинальные – диаминопимелиновая кислота, D-аланин, D-глютанин, входящие в состав пептидогликанов и капсул некоторых бактерий. Только в спорах находится дипиколиновая кислота, с которой связана высокая резистентность спор. Жгутики построены из белка флагеллина, обладающего сократительной способностью и выраженными антигенными свойствами. Пили (ворсинки) содержат особый белок - пилин. Пептидную природу имеют капсулы представителей рода Bacillus, возбудителя чумы, поверхностные антигены ряда бактерий, в том числе стафилококков и стрептококков. Белок А - специфический белок S.aureus - фактор, обуславливающий ряд свойств этого возбудителя. Белок М - специфический белок гемолитических стрептококков серогруппы А, позволяющий дифференцировать серовары (около 100), что имеет эпидемиологическое значение. Ряд белков содержит наружная мембрана грамотрицательных бактерий, из которых 3 - 4 мажорных (основных) и более 10- второстепенных, выполняющих различные функции. Среди мажорных белков - порины, образующие диффузные поры, через которые в клетку могут проникать мелкие гидрофильные молекулы. Белки входят в состав пептидогликана - биополимера, составляющего основу бактериальной клеточной стенки. Он состоит из остова (чередующиеся молекулы двух аминосахаров) и двух наборов пептидных цепочек - боковых и поперечных. Наличие двух типов связей - гликозидных (между аминосахарами) и пептидных, которые соединяют субъединицы пептидогликанов, придают этому гетерополимеру структуру молекулярной сети. Пептидогликан - наиболее устойчивое соединение, которое образует ригидную мешковидную макромолекулу, определяющую постоянную форму бактерий и ряд их свойств. 1.Пептидогликан обладает антигенными свойствами (содержит родо - и видоспецифические антигенные детерминанты). 2.Он запускает классический и альтернативный пути активации системы комплемента. 3.Пептидогликан тормозит фагоцитарную активность и миграцию макрофагов. 4.Он способен инициировать развитие гиперчувствительности замедленного типа (ГЗТ). 5.Пептидогликан обладает противоопухолевым действием. 6.Он оказывает пирогенное действие, т.е. вызывает лихорадку. Из соединений белков с небелковыми компонентами наибольшее значение имеют липопротеиды, гликопротеиды и нуклеопротеиды. Удивительное таинство жизни - синтез белка осуществляется в рибосомах. Существует два основных типа рибосом - 70S (S- константа седиментации, единица Сведберга) и 80S. Рибосомы первого типа встречаются только у прокариотов. Антибиотики не действуют на синтез белка в рибосомах типа 80S, распространенных у эукариотов. Липиды (главным образом форфолипиды) содержатся в цитоплазматической мембране (липидный бислой), в также в наружной мембране грамотрицательных бактерий. Есть микроорганизмы, содержащие большое количество липидов (до 40% сухого остатка)- микобактерии. В состав липидов входят различные жирные кислоты, весьма специфичные для разных групп микроорганизмов. Их определение имеет в ряде случаев диагностическое значение, например у анаэробов, микобактерий. У микобактерий туберкулеза в составе липидов имеется ряд кислотоустойчивых жирных кислот - фтионовая, миколовая и др. Высокое содержание липидов и их состав определяют многие свойства микобактерий туберкулеза: -устойчивость к кислотам, щелочам и спиртам; -трудная окрашиваемость красителями (используют специальные методы окраски, чаще - по Цилю - Нильсену); -устойчивость возбудителя к солнечной радиации и дезосредствам; - патогенность. Тейхоевые кислоты встречаются в клеточных стенках грамположительных бактерий. Представляют собой водорастворимые линейные полимеры, содержащие остатки глицерина или рибола, связанные фосфодиэфирными связями. С тейхоевыми кислотами связаны главные поверхностные антигены ряда грамположительных бактерий. Углеводы встречаются чаще в виде полисахаридов, которые могут быть экзо - и эндоклеточными. Среди экзоклеточных полисахаридов выделяют каркасные (входят в состав капсул) и истинно экзополисахариды (выходят во внешнюю среду). Среди бактериальных полисахаридов многие находят медицинское применение. Декстраны - полисахариды с большой молекулярной массой, по виду напоминают слизь. 6% раствор - кровезаменитель полиглюкин. Декстрановый гель сефадекс используется в колоночной хроматографии как молекулярное сито. Эндоклеточные полисахариды - запасные питательные вещества клетки (крахмал, гликоген и др.). Липополисахарид (ЛПС) - один из основных компонентов клеточной стенки грамотрицательных бактерий, это соединение липида с полисахаридом. ЛПС состоит из комплекса: 1.Липид А. 2.Одинаковое для всех грамотрицательных бактерий полисахаридное ядро. 3.Терминальная сахаридная цепочка (О - специфическая боковая цепь). Синонимы ЛПС - эндотоксин, О - антиген. ЛПС выполняет две основные функции - определяет антигенную специфичность и является одним из основных факторов патогенности. Это- эндотоксин, токсические свойства которого проявляются преимущественно при разрушении бактериальных клеток. Его токсичность определяется липидом А. ЛПС запускает синтез более 20 биологически активных веществ, определяющих патогенез эндотоксикоза, обладает пирогенным действием. Нуклеиновые кислоты - ДНК и РНК. Рибонуклеиновые кислоты (РНК) находятся главным образом в рибосомах (р-РНК- 80- 85%), т (транспортные)- РНК- 10%, м (матричные)- РНК- 1- 2%, главным образом в одноцепочечной форме. ДНК (дезоксирибонуклеиновая кислота) может находиться в ядерном аппарате (хромосомная ДНК) или в цитоплазме в специализированных образованиях - плазмидах - плазмидная (внехромосомная) ДНК. Микроорганизмы отличаются по структуре нуклеиновых кислот, содержанию азотистых оснований. Генетический код состоит всего из четырех букв (азотистых оснований) - А (аденин), Т (тимин), Г (гуанин) и Ц (цитозин). Наиболее часто для характеристики микроорганизмов используют как таксономический признак процентное соотношение Г/Ц, которое существенно отличается у различных групп микроорганизмов. 20.Ферментыбактерий. Основныеклассы,генетическийконтроль, классификация,характеристикаферментов вирулентности. Микроорганизмы синтезируют различные ферменты - специфические белковые катализаторы. У бактерий обнаружены ферменты 6 основных классов. 1.Оксидоредуктазы - катализируют окислительно-восстановительные реакции. 2.Трансферазы - осуществляют реакции переноса групп атомов. 3.Гидролазы - осуществляют гидролитическое расщепление различных соединений. 4.Лиазы - катализируют реакции отщепления от субстрата химической группы негидролитическим путем с образованием двойной связи или присоединения химической группы к двойным связям. 5.Лигазы или синтетазы - обеспечивают соединение двух молекул, сопряженное с расщеплением пирофосфатной связи в молекуле АТФ или аналогичного трифосфата. 6.Изомеразы - определяют пространственное расположение групп элементов. В соответствии с механизмами генетического контроля у бактерий выделяют три группы ферментов: - конститутивные, синтез которых происходит постоянно; - индуцибельные, синтез которых индуцируется наличием субстрата; - репрессибельные, синтез которых подавляется избытком продукта реакции. Ферменты бактерий делят на экзо - и эндоферменты. Экзоферменты выделяются во внешнюю среду, осуществляют процессы расщепления высокомолекулярных органических соединений. Способность к образованию экзоферментов во многом определяет инвазивность бактерий - способность проникать через слизистые, соединительнотканные и другие тканевые барьеры. Примеры: гиалуронидаза расщепляет гиалуроновую кислоту, входящую в состав межклеточного вещества, что повышает проницаемость тканей (клостридии, стрептококки, стафилококки и многие другие микроорганизмы); нейраминидаза облегчает преодоление слоя слизи, проникновение внутрь клеток и распространение в межклеточном пространстве (холерный вибрион, дифтерийная палочка, вирус гриппа и многие другие). К этой же группе относятся энзимы, разлагающие антибиотики. В бактериологии для дифференциации микроорганизмов по биохимическим свойствам основное значение часто имеют конечные продукты и результаты действия ферментов. В соответствии с этим существует микробиологическая (рабочая) классификация ферментов. 1.Сахаролитические. 2.Протеолитические. 3.Аутолитические. 4.Окислительно-восстановительные. 5.Ферменты патогенности (вирулентности). Ферментный состав клетки определяется геномом и является достаточно постоянным признаком. Знание биохимических свойств микроорганизмов позволяет идентифицировать их по набору ферментов. Основные продукты ферментирования углеводов и белков - кислота, газ, индол, сероводород, хотя реальный спектр для различных микроорганизмов намного более обширный. Основные ферменты вирулентности - гиалуронидаза, плазмокоагулаза, лецитиназа, нейраминидаза, ДНК-аза. Определение ферментов патогенности имеет значение при идентификации ряда микроорганизмов и выявления их роли в патологии. Ряд ферментов микроорганизмов широко используется в медицине и биологии для получения различных веществ (аутолитические, протеолитические), в генной инженерии (рестриктазы, ДНК - лигазы, ДНК - полимеразы, обратная транскриптаза). Дополнение: Ферменты (от греч. fermentum ― закваска ) ― высокоспецифические белковые катализаторы, присутствующие во всех живых клетках, без которых не возможны жизнь и размножение. Ферменты распознают соответствующие им метаболиты (субстраты), вступают с ними во взаимодействие и ускоряют химические реакции. Ферменты являются белками. Ферментный состав микроорганизма определяется геномом и является достаточно стабильным признаком. Определение ферментов широко применяется для биохимической идентификации бактерий. Эндоферменты катализируют метаболизм проходящий внутри клетки. Экзоферменты выделяются клеткой в окружающую среду. Конститутивные ферменты постоянно синтезируются в определенных концентрациях. Индуцибельные ферменты – это ферменты, концентрация которых увеличивается при поступлении соответствующего субстрата. Ферменты агрессии: гиалуронидаза, фибринолизин, нейраминидаза, коллагеназа, лецитиназа (лецитовителлаза), коагулаза, уреаза, аминокислотные декарбоксилазы, дезоксирибонуклеаза. 21.Метаболизм микроорганизмов. Механизмы поступления веществ. Классификация исостав питательныхсред. Метаболизм бактерий― совокупность физико-химических процессов (химических превращений и реакций), направленных на воспроизводство структур и обеспечение жизненных функций микробной клетки, таких как: рост и размножение; отложение резервного пищевого материала; транспорт питательных веществ в микробную клетку; выделение продуктов метаболизма (токсинов, ферментов, антибиотиков и других биологически активных веществ); движение; спорообразование; адгезия на рецепторах клеток хозяина и проникновение в них; • различных адаптивных реакций на изменение внешней среды. Анаболизм — совокупность биохимических реакций, осуществляющих синтез компонентов клетки. Катаболизм — совокупность реакций, обеспечивающих клетку энергией. Схема изучения метаболизма – этапы: Начальный (периферический) метаболизм – проникновение веществ в клетку извне и распад до промежуточных продуктов. Амфиболизм (промежуточный метаболизм) – образование промежуточных продуктов метаболизма, общих для катаболических и анаболических путей. Конечные, строго специализированные этапы конструктивного метаболизма (ведут к построению структур клетки) и энергетического метаболизма (образование АТФ). Механизмы проникновения питательных веществ в клетку: Простая диффузия (для истинных растворов). Энергонезависимый процесс. Облегченная диффузия («паром по течению») – в направлении градиента концентрации с участием белков – переносчиков. Энергозависимый процесс. Активный транспорт – против концентрационного и электрохимического градиента с участием пермеаз (амино-, оксикислотных, ионных и др.). Процесс идет с затратой энергии АТФ, зависит от заряда веществ и их трансформации в процессе переноса. Микроорганизмы по способности усваивать источники углерода делятся на две группы: автотрофы (лат. autos — сам, trophe — питание) синтезируют все углеродсодержащие компоненты клетки из СО2 как единственного источника углерода и гетеротрофы (лат. heteros — другой, «питающийся за счет других») используют разнообразные органические углеродсодержащие соединения. В зависимости от источников энергии и микроорганизмы подразделяют на фототрофы (фотосинтезирующие), способные использовать солнечную энергию, и хемотрофы (хемосинтезирующие), получающие энергию за счет окислительно-восстановительных реакций. В зависимости от используемых доноров электронов бактерии разделяют на литотрофы (используют неорганические доноры электронов) и органотрофы (используют органические соединения). Прототрофы ― микроорганизмы, способные синтезировать все необходимые им органические соединения из глюкозы и солей аммония. Ауксотрофы ― микроорганизмы, не способные синтезировать какие-либо органические соединения. Они получают эти соединения в готовом виде из окружающей среды или организма человека. Классификация питательных сред По консистенции: жидкие, полужидкие, плотные. По происхождению: естественные (молоко, картофель), искусственные, полусинтетические, синтетические. По составу: простые (МПА, МПБ, овощи, молоко), сложные (1% глюкозы, 10−20% сыворотки крови, 20−30% асцитической жидкости, 5−10% дефибринированной крови). По назначению: универсальные — среды, на которых хорошо растут многие виды бактерий. К ним относятся мясо-пептонный бульон (МПБ) и мясопептонный агар (МПА); специальные ― среды, специально приготовленные для получения роста бактерий, которые не растут на универсальных средах; дифференциально-диагностические ― среды, позволяющие отличать одни виды бактерий от других по ферментативной активности; селективные — среды, содержащие вещества, используемые микроорганизмами определенных видов и препятствующие росту других микроорганизмов. Селективные среды позволяют направленно отбирать из исследуемого материала определенные виды бактерий; дифференциально-селективные — среды, сочетающие в себе свойства дифференциально-диагностических и селективных сред; • консервирующие; обогатительные.  Дополнение: Метаболизм микроорганизмов. Для роста и размножения микроорганизмы нуждаются в веществах, используемых для построения структурных компонентов клетки и получения энергии. Метаболизм (т.е. обмен веществ и энергии) имеет две составляющих - анаболизм и катаболизм. Анаболизм- синтез компонентов клетки (конструктивный обмен). Катаболизм - энергетический обмен, связан с окислительно-восстановительными реакциями, расщеплением глюкозы и других органических соединений, синтезом АТФ. Питательные вещества могут поступать в клетку в растворимом виде (это характерно для прокариотов) - осмотрофы, или в виде отдельных частиц - фаготрофы. Основным регулятором поступления веществ в бактериальную клетку является цитоплазматическая мембрана. Существует четыре основных механизма поступления веществ: - пассивная диффузия - по градиенту концентрации, энергонезатратная, не имеющая субстратной специфичности; - облегченная диффузия - по градиенту концентрации, субстратспецифичная, энергонезатратная, осуществляется при участии специализированных белков пермеаз; - активный транспорт - против градиента концентрации, субстратспецифичен (специальные связывающие белки в комплексе с пермеазами), энергозатратный (за счет АТФ), вещества поступают в клетку в химически неизмененном виде; - транслокация (перенос групп) - против градиента концентрации, с помощью фосфотрансферазной системы, энергозатратна, вещества (преимущественно сахара) поступают в клетку в форфорилированном виде. Основные химические элементы - органогены, необходимые для синтеза органических соединений - углерод, азот, водород, кислород. В зависимости от источника потребляемого углерода микробы подразделяют на аутотрофы (используют CO2) и гетеротрофы (используют готовые органические соединения). В зависимости от источника энергии микроорганизмы делят на фототрофы (энергию получают за счет фотосинтеза - например, цианобактерии) и хемотрофы (энергия добывается за счет химических, окислительно-восстановительных реакций). Если при этом донорами электронов являются неорганические соединения, то это литотрофы, если органические - органотрофы. Если бактериальная клетка в состоянии синтезировать все необходимые для жизнедеятельности вещества, то это прототрофы. Если бактерии нуждаются в дополнительных веществах (факторах роста), то это ауксотрофы. Основными факторами роста для трудно культивируемых бактерий являются пуриновые и пиримидиновые основания, витамины, некоторые (обычно незаменимые) аминокислоты, кровяные факторы (гемин) и др. Краткая характеристика питательных сред. По консистенции выделяют жидкие, плотные (1,5- 3% агара) и полужидкие (0,3- 0,7 % агара) среды. Агар- полисахарид сложного состава из морских водорослей, основной отвердитель для плотных (твердых) сред. В качестве универсального источника углерода и азота применяют пептоны- продукты ферментации белков пепсином, различные гидролизаты- мясной, рыбный, казеиновый, дрожжевой и др. По назначению среды разделяют на ряд групп: - универсальные (простые), пригодные для различных нетребовательных микроорганизмов (мясо - пептонный бульон - МПБ, мясо - пептонный агар - МПА); - специальные - среды для микроорганизмов, не растущих на универсальных средах (среда Мак - Коя на туляремию, среда Левенштейна- Иенсена для возбудителя туберкулеза); - дифференциально- диагностические - для дифференциации микроорганизмов по ферментативной активности и культуральным свойствам (среды Эндо, Плоскирева, Левина, Гисса); - селективные и элективные - для выделения определенных видов микроорганизмов и подавления роста сопутствующих - пептонная вода, селенитовая среда, среда Мюллера. По происхождению среды делят на естественные, полусинтетические и синтетические. 22.Типы дыхания микроорганизмов. Методысоздания анаэробныхусловий для культивирования. Основныепринципыкультивирования. Путем дыхания микроорганизмы добывают энергию. Дыхание - биологический процесс переноса электронов через дыхательную цепь от доноров к акцепторам с образованием АТФ. В зависимости от конечного акцептора электронов выделяют аэробное и анаэробное дыхание. При аэробном дыхании конечным акцептором электронов является молекулярный кислород (О2), при анаэробном - связанный кислород ( -NO3 , =SO4, =SO3). По типу дыхания выделяют четыре группы микроорганизмов. 1.Облигатные (строгие) аэробы. Им необходим молекулярный (атмосферный) кислород для дыхания. 2.Микроаэрофилы нуждаются в уменьшенной концентрации (низком парциальном давлении) свободного кислорода. Для создания этих условий в газовую смесь для культивирования обычно добавляют CO2, например до 10- процентной концентрации. 3.Факультативные анаэробы могут потреблять глюкозу и размножаться в аэробных и анаэробных условиях. Среди них имеются микроорганизмы, толерантные к относительно высоким (близких к атмосферным) концентрациям молекулярного кислорода - т.е. аэротолерантные, а также микроорганизмы, которые способны в определенных условиях переключаться с анаэробного на аэробное дыхание. 4.Строгие анаэробы размножаются только в анаэробных условиях, т.е. при очень низких концентрациях молекулярного кислорода, который в больших концентрациях для них губителен. Биохимически анаэробное дыхание протекает по типу бродильных процессов, молекулярный кислород при этом не используется. Аэробное дыхание энергетически более эффективно (синтезируется большее количество АТФ). В процессе аэробного дыхания образуются токсические продукты окисления (H2O2- перекись водорода, -О2 - свободные кислородные радикалы), от которых защищают специфические ферменты, прежде всего каталаза, пероксидаза, пероксиддисмутаза. У анаэробов эти ферменты отсутствуют, также как и система регуляции окислительно-восстановительного потенциала (rH2). Основные методы создания анаэробных условий для культивирования микроорганизмов. 1.Физические методы - откачивание воздуха, введение специальной газовой безкислородной смеси (чаще- N2- 85%, CO2- 10%, H2- 5%). 2.Химические методы - применяют химические поглотители кислорода. 3.Биологические методы - совместное культивирование строгих аэробов и анаэробов (аэробы поглощают кислород и создают условия для размножения анаэробов). 4.Смешанные методы - используют несколько разных подходов. Необходимо отметить, что создание оптимальных условий для строгих анаэробов - очень сложная задача. Очень непросто обеспечить постоянное поддержание безкислородных условий культивирования, необходимы специальные среды без содержания растворенного кислорода, поддержание необходимого окислительно-восстановительного потенциала питательных сред, взятие и доставка, посев материала в анаэробных условиях. Существует ряд приемов, обеспечивающих более подходящие условия для анаэробов - предварительное кипячение питательных сред, посев в глубокий столбик агара, заливка сред вазелиновым маслом для сокращения доступа кислорода, использование герметически закрывающихся флаконов и пробирок, шприцев и лабораторной посуды с инертным газом, использование плотно закрывающихся эксикаторов с горящей свечой. Используются специальные приборы для создания анаэробных условий - анаэростаты. Однако в настоящее время наиболее простым и эффективным оборудованием для создания анаэробных и микроаэрофильных условий является система “Газпак” со специальными газорегенерирующими пакетами, действующими по принципу вытеснения атмосферного воздуха газовыми смесями в герметически закрытых емкостях. Основные условия культивирования микроорганизмов на питательных средах. 1.Использование всех необходимых для соответствующих микробов питательных компонентов. 2.Оптимальные температура, рН, rH2, концентрация ионов, степень насыщения кислородом, газовый состав и давление. Микроорганизмы культивируют на питательных средах при оптимальной температуре в термостатах, обеспечивающих условия инкубации. По температурному оптимуму роста выделяют три основные группы микроорганизмов. 1.Психрофилы - растут при температурах ниже +20 градусов Цельсия. 2.Мезофилы - растут в диапозоне температур от 20 до 45 градусов (часто оптимум - при 37 градусах С). 3.Термофилы - растут при температурах выше плюс 45 градусов. 23.Рост иразмножениемикроорганизмов. Периодические, непрерывныеи синхронные культуры. Рост и размножение микроорганизмов. Бактериальные клетки размножаются в результате деления. Основные стадии размножения микробов в жидкой среде в стационарных условиях: - лаг- фаза (начальная стадия адаптации с медленным темпом прироста биомассы бактерий); - экспоненциальная (геометрического роста) фаза с резким ростом численности популяции микроорганизмов (2 в степени n); - стационарная фаза (фаза равновесия размножения и гибели микробных клеток); - стадия гибели - уменьшение численности популяции в связи с уменьшением и отсутствием условий для размножения микроорганизмов (дефицит питательных веществ, изменение рH, rH2, концентрации ионов и других условий культивирования). Данная динамика характерна для периодических культур с постепенным истощением запаса питательных веществ и накоплением метаболитов. Если в питательной среде создают условия для поддержания микробной популяции в экспоненциальной фазе - это хемостатные (непрерывные) культуры. Характер роста бактерий на плотных и жидких питательных средах: сплошной рост, образование колоний, осадок, пленка, помутнение. На плотных средах отмечается сплошной рост или образование колоний. Колонии можно описать рядом характеристик - форма, размер, поверхность, профиль, прозрачность, цвет, край, структура, консистенция. На жидких средах наблюдают помутнение (чаще - факультативные анаэробы), поверхностный рост в виде пленок (аэробные прокариоты), пристеночный, придонный рост, образования различных по характеристикам осадков. Чистая культура - популяция одного вида микроорганизмов. Основные принципы получения чистых культур: механическое разобщение, рассев, серийные разведения, использование элективных сред, особых условий культивирования (с учетом устойчивости некоторых микробов к определенным температурам, кислотам, щелочам, парциальному давлению кислорода, рН и мн. др). Классические методы - Пастера (разведения в жидкой среде), Коха (пластинчатые разводки), Шукевича (ползучий рост), Дригальского (посев одним шпателем последовательно в три чашки Петри), Вейнберга (для анаэробов - заливают агаровую среду сверху смесью парафина и вазелинового масла). В лабораторных и промышленных условиях используют два основных способа культивирования микроорганизмов: периодическое и непрерывное. Периодическая культура - это популяция клеток в ограниченном жизненном пространстве. Рост бактерий в периодической культуре происходит до тех пор, пока содержание какого-нибудь из необходимых им компонентов питательной среды не достигнет минимума, после чего рост прекращается. Зависимость концентрации жизнеспособных клеток при периодическом культивировании от длительности инкубирования описывается характерной кривой, которая имеет S-образную форму (рис. 1). На кривой можно различить несколько фаз роста, сменяющих друг друга в определенной последовательности: лаг-фазу; логарифмическую фазу; стационарную фазу; фазу отмирания. 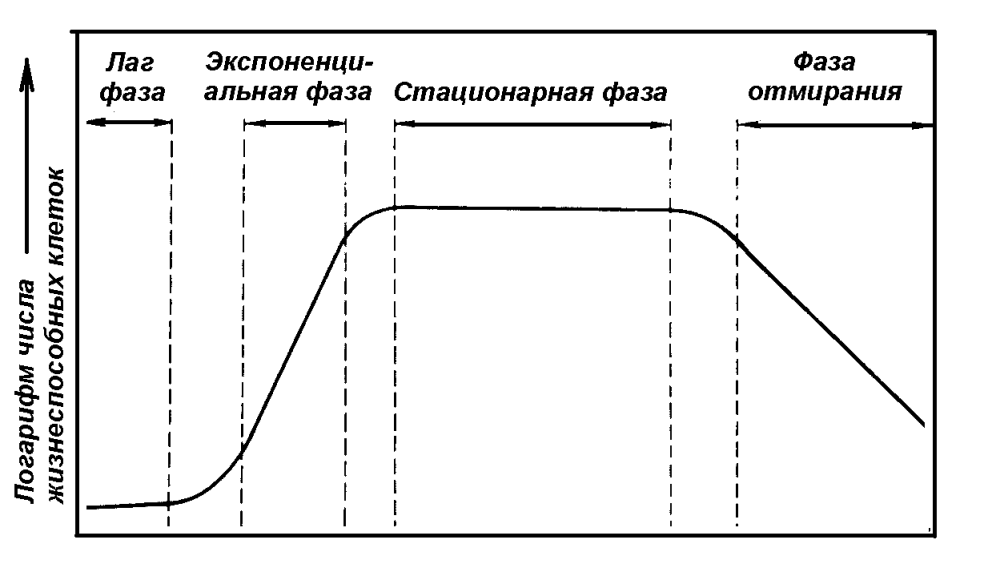 Рис. 1. Основные фазы кривой роста периодической культуры микроорганизмов В условиях непрерывного культивирования в сосуд, содержащий популяцию бактерий, подается свежая питательная среда и из него одновременно удаляется часть среды с клетками микроорганизмов. Это позволяет на длительное время задержать культуру в состоянии логарифмического роста. Для проточного глубинного культивирования бактерий с аэрацией в промышленных и лабораторных условиях применяют биореакторы, или ферментеры. Они представляют собой герметические котлы, в которые заливается жидкая питательная среда. Ферментеры снабжены автоматическими приспособлениями, позволяющими поддерживать постоянную температуру, оптимальное значение рН и редокс-потенциал, дозированное поступление необходимых питательных веществ и т.д. Кроме того, они снабжены системами перемешивания, аэрирования, охлаждения, пеногашения. Ферментеры бывают нескольких типов: хемостаты, турбидостаты, оксистаты, рН-статы и др. (т.е. различаются по способу управления). Хемостат состоит из сосуда-культиватора, в который с заданной постоянной скоростью поступает питательная среда. Для равномерного и полного смешивания питательных веществ содержимое культиватора механически перемешивается и аэрируется стерильным воздухом. Избыточная биомасса клеток с питательной средой вытекает из культиватора через сливной сифон. В хемостате прирост биомассы прямо пропорционален скорости притока субстрата и удаления продуктов метаболизма. Примером хемостата в природе служит рубец жвачных животных. Турбидостат представляет собой ферментер, в котором поддерживается заданная плотность клеток за счет определения оптической плотности среды культивирования. Когда количество биомассы увеличивается относительно некоторого выбранного уровня, что фиксируется фотоэлементом, соединенным с системой реле, включается подача свежей питательной среды. Непрерывное культивирование широко используется в промышленной микробиологии, а также при проведении физиологических, биохимических и генетических исследований, так как в данных условиях наблюдается постоянная плотность популяции и концентрации всех компонентов питательной среды. |