контрольная работа. готовая контрольная работа по биохимии. Аминокислоты это строительные блоки макромолекул белков. По строению они являются органическими карбоновыми кислотами, у которых, как минимум, один атом водорода замещен на аминогруппу. Классификация
 Скачать 209.56 Kb. Скачать 209.56 Kb.
|

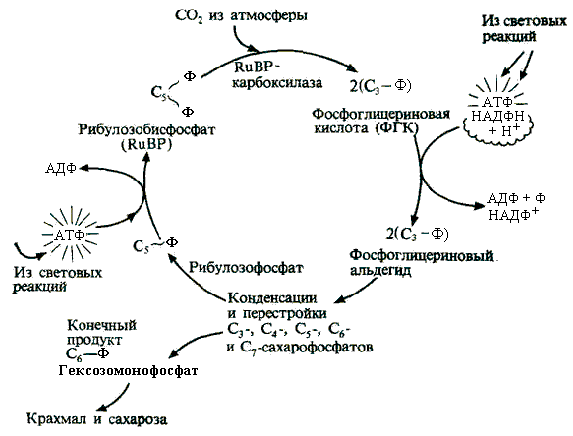
10 % энергии растрачивается на поддержание цикла.Контрольная работа Вариант 10 Аминокислоты – это строительные блоки макромолекул – белков. По строению они являются органическими карбоновыми кислотами, у которых, как минимум, один атом водорода замещен на аминогруппу. КЛАССИФИКАЦИЯ 1. В зависимости от положения аминогруппы Выделяют α, β, γ и другие аминокислоты. Для организма млекопитающих наиболее характерны α- аминокислоты. 2. Для медиков наиболее интересна классификация для оп участию аминокислот в синтезе белков: протеиногенные (20 АК) и непротеиногенные (около 40 АК). 3. По абсолютной конфигурации молекулы – D- и L-формы. Различия связаны со взаимным расположением четырех замещающих групп, находящихся в вершинах воображаемого тетраэдра. Как известно, тетраэдр – это довольно жесткая структура, в которой невозможно произвольным образом передвинуть вершины. 4. По оптической активности – право- и левовращающие. Наличие ассиметричного атома С (хирального центра) делает возможным только два расположения химических групп вокруг него. Это приводит к особому отличию веществ друг от друга, а именно – изменению направления вращения плоскости поляризации поляризованного света, проходящего через раствор. Величину угла поворота определяют при помощи поляриметра. В соответствии вращающие (+) и левовращающие (–) изомеры В растениях реакции протекают с большой скоростью, потому что в любой живой клетке находятся многочисленные биологические катализаторы, называемые ФЕРМЕНТАМИ или ЭНЗИМАМИ, которые способны в тысячи и даже миллионы раз ускорять течение химических реакций. ФЕРМЕНТЫ - специфические катализаторы белковой природы, ускоряющие течение определенных химических реакций и играющие важнейшую роль в обмене веществ. Механизм действия ферментов как раз и заключается в снижении энергии активации, необходимой для прохождения химической реакции. Ферментативная реакция осуществляется в несколько этапов 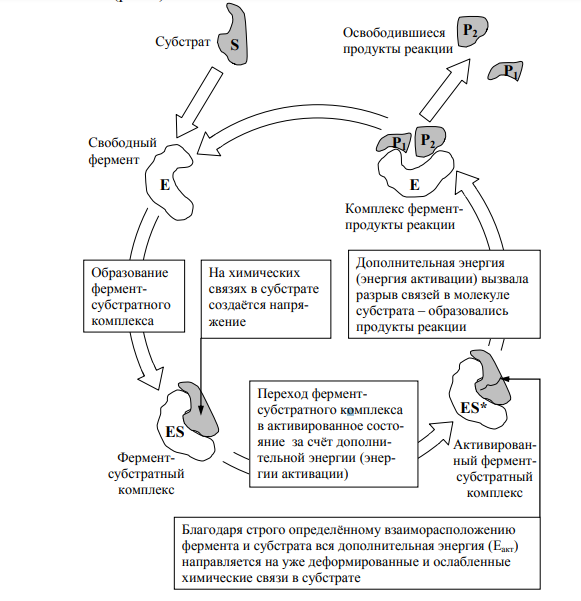 Фермент-субстратный комплекс за счет дополнительной энергии, называемой энергией активации, переходит в активированное состояние – активированный комплекс ES*. Дополнительная энергия (энергия активации) обеспечивает разрыв определенных химических связей в субстрате и образование комплекс продуктов реакции с ферментом (ЕР). ДИНУКЛЕОТИДЫ Состоят из двух нуклеотидов, но имеют особенности в строении. Наиболее известными являются: никотинамидадениндинуклеотид (НАД+), никотинамидадениндинуклеотидфосфат (НАДФ+). Главной функцией является перенесение электронов (2) и иона водорода (1). Могут восстанавливаться: НАД+ + 2е— + Н+ →НАДН; НАДФ+ + 2е— + Н+ →НАДФН. На определенном участке для некоторых реакций эти соединения отдают протон водорода, электроны: НАДН →НАД+ + 2е— + Н+; НАДФН → НАДФ+ + 2е— + Н+ Темновая фаза фотосинтеза Темновая фаза фотосинтеза – это комплекс ферментативных реакций, во время которой происходит восстановление поглощенного углекислого газа за счет продуктов световой фазы (АТФ и НАДФН). Различают несколько циклов восстановления СО2. Цикл Кальвина. Этот способ ассимиляции СО2 является основным и присущ всем растениям. Он был расшифрован американскими учеными во главе с М. Кальвином. В 1961 году М. Кальвину за установление последовательности реакций в этом цикле и была присуждена Нобелевская премия. Этот цикл начинается с присоединения СО2 к акцептору – пятиуглеродному сахару рибулозо-1,5-дифосфату (РДФ). Присоединение СО2 к тому или ионному веществу называется карбоксилированием, а фермент катализирующий такую реакцию – карбоксилазой. В данной реакции карбоксилирование происходит с участием фермента рибулозодифосфаткарбоксилазы (РДФ-карбоксилаза). Это самый распространенный в мире фермент. Продукт реакции, содержащий 6 атомов углерода, в присутствии воды сразу распадается на две молекулы 3-фосфоглицириновой кислоты (3-ФГК): 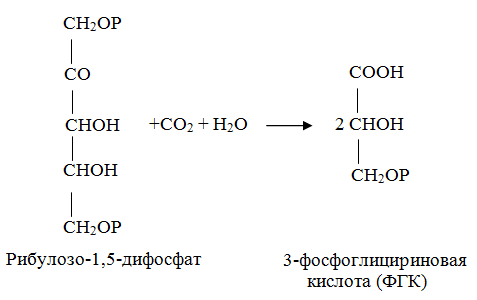 С данной реакции и начинается цикл Кальвина. ФГК и является, по современным взглядам, первичным продуктом ассимиляции углерода. Для дальнейших превращений ФГК необходимы вещества световой фазы фотосинтеза: АТФ и НАДФН. Сначала 3-ФГК фосфорилируется при участии АТФ и образуется 1,3-дифосфоглицириновая кислота. Реакция катализируется ферментом фосфоглицераткиназой: 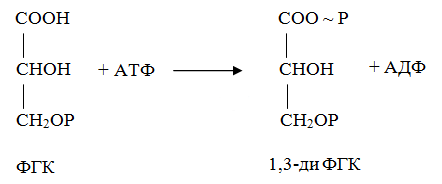 атем происходит восстановление за счет НАДФН и образуется фосфоглицириновый альдегид ФГА: 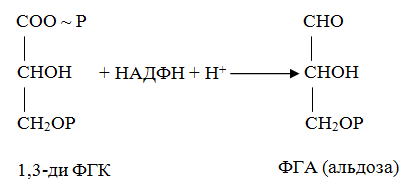 Суммарный результат второй стадии: восстановление карбоксильной группы кислоты (–СООН) до альдегидной (–СНО). Процесс превращения катализируется дегидрогеназой фосфоглициринового альдегида. Дальнейшее превращение фосфоглициринового альдегида может происходить 4 путями. ФГА частично с помощью триозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА): 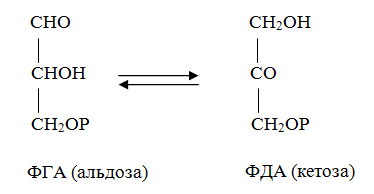 Это первый путь превращения ФГА. Таким образом, в клетку поступают две найпростейшие формы сахаров: альдоза (ФГА) и кетоза (ФДА). Это трехуглеродные сахара (триозосахара) с присоединенной к ним фосфатной группой содержат больше химической энергии, чем ФГК. Это первые углеводы, которые образуются при фотосинтезе. С помощью альдолазы фосфодиоксиацетон (ФДА) соединяется с другой молекулой ФГА и образуется молекула фруктозо-1,6-дифосфата (ФДФ). 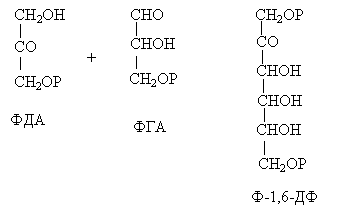 Это второй путь превращения ФГА. Фруктозо-1,6-дифосфат дефосфорилируется и превращается во фруктозо-6-фосфат (Ф-6-Ф), что сопровождается накоплением в среде неорганического фосфата. Фруктозо-6-фосфат в дальнейшем может выйти из цикла и использоваться для синтеза запасных форм углеводов: сахарозы, крахмала, других полисахаридов. Однако ФГА (третий путь) может реагировать с эквимолярным количеством Ф-6-Ф, в результате образуются равные количества ксилулозо-5-фосфата и эритрозо-4-фосфата (транскетолаза). Затем эритрозо-4-фосфат реагирует с равным количеством ФДА и образуется седагептулозо-1,7-дифосфат (альдолаза), которая фосфорилируется до седагептулозо-7-фосфата с участием седагептулозодифосфатазы. 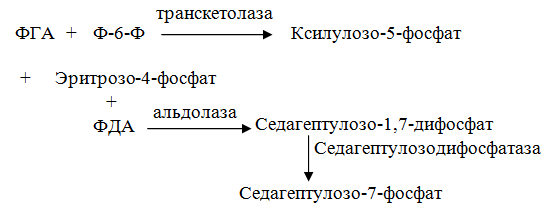 Четвертый путь превращения ФГА связан с его реакцией с седагептулозо-7-фосфатом с образованием равных (эквимолярных) количеств рибозо-5-фосфата и ксилулозо-5-фосфата. Ксилулозо-5-фосфат эпимиризуется, а рибозо-5-фосфат изомерезуется до рибулозо-5-фосфата, последняя фосфорилируется за счет АТФ и образуется рибулозо-1,5-дифосфат – первичное соединение цикла Кальвина (акцептор СО2). В этих реакциях тратится еще три молекулы АТФ. 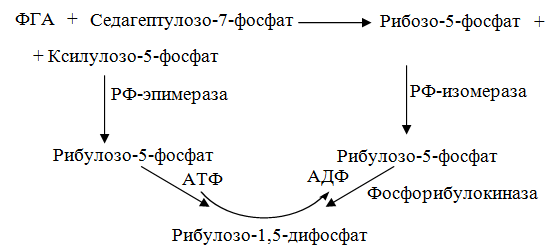 Из приведенных реакций цикла Кальвина видно, что фотосинтез, являясь процессом запасания энергии, тем не менее, для своего существования требует затраты энергии. В цикле Кальвина образование фруктозо-6-фосфата можно представить в виде следующего суммарного выражения: 6СО2 + 12НАДФН + 12Н+ + 18АТФ + 11Н2О → фруктозо-6-фосфат + 12НАДФ+ + 18АДФ + 17Фн 18 молекул АТФ запасают около 140 ккал и 12 НАДФН – |