|
|
Раздел 1. Микробиология. 1. Предмет и з
Синхронные культуры – это популяции микроорганизмов, в которых все клетки находятся на одинаковой стадии клеточного цикла и делятся одновременно.Такие культуры часто необходимы для изучения процессов обмена веществ на протяжении цикла клеточного деления.
Синхронизировать рост и деление клеток в какой-либо популяции можно с использованием различных приемов, таких как: изменение температуры; изменение интенсивности освещения (для фототрофных микроорганизмов); лимитирование количества питательных веществ (т.н. «метаболический шок» - когда сначала выращивают бактерии на голодном пайке, а потом вносят в среду необходимые питательные вещества); фильтрование суспензии клеток микроорганизмов через специальный фильтр, позволяющий отобрать клетки одного размера; центрифугирование. Несмотря на разнообразие методов, которые могут привести культуру к синхронному размножению, не всегда удается в достаточной степени синхронизировать деление клеток. В таких случаях принято сочетать различные методы. Однако в синхронизированном состоянии культура не может находиться длительное время и после двух-трех генераций процесс деления клеток асинхронизируется.
Синхронизации деления подвергались различные объекты: грибы, бактерии, простейшие, водоросли, а также клетки культуры тканей растений и животных.
Дополнение:
Размножение бактерий на жидких и плотных питательных средах.
Рост ―координированное воспроизведение всех компонентов бактериальной клетки и увеличение ее биомассы. Размножение ― воспроизводство и увеличение количества клеток, приводящее к образованию бактериальной популяции.
Бактерии характеризуются высокой скоростью размножения. Скорость размножения зависит от видовой принадлежности, состава питательной среды, рН, температуры, аэрации.
На плотных питательных средах бактерии образуют скопления клеток, называемые колониями. Колонии разных видов отличаются по размерам, форме, консистенции, окраске, характеру краев, характеру поверхности, прозрачности. Характер роста на жидких питательных средах: пленочный (образованием пленки на поверхности питательной среды), диффузное помутнение, придонный (образование осадка).
Фазы развития бактериальной популяции
Исходная стационарная фаза (
1−2 ч.). Число бактерий не увеличивается, клетки не растут.
Лаг-фаза или фаза задержки размножения ( 2 ч.).
Log-фаза ― логарифмическая или экспоненциальная фаза ( 3−5 ч). Популяция делится с максимальной скоростью и идет увеличение особей в геометрической прогрессии.
Фаза отрицательного ускорения ( 2 ч.). Связана с истощением лимитирующего метаболита или накоплением токсических продуктов метаболизма.
Стационарная фаза максимума. Количество образующихся и отмирающих клеток одинаково.
Фаза ускоренной гибели ( 3 ч.).
Логарифмическая фаза гибели ( 5 ч.).
Фаза уменьшения скорости отмирания – остающиеся живые особи переходят в состояние покоя.
24.Классификация, структура и особенности биологии вирусов.
Предмет изучения раздела медицинской вирусологии - эпидемиология, патогенез, клиника заболеваний, вызываемых вирусами, а также методы лабораторной диагностики, специфической профилактики и лечения вирусных инфекций.
Вирусы - это мельчайшие формы жизни, не имеющие клеточного строения. Вирусы образуют отдельное царство (Vira). От других микробов вирусы отличаются присущими только им уникальными свойствами.
Ультрамикроскопические размеры вирусов. Вирусы измеряются в нанометрах (1 мм = 1000 мкм, 1 мкм = 1000 нм). По размерам вирусы подразделяются на мелкие (например, вирус полиомиелита) - размер вирусных частиц 10-25 нм, средние (например, вирусы гриппа) - размер вирионов составляет 100-120 нм, крупные (например, вирус натуральной оспы) - размер вирусных частиц около 350 нм.
Наличие только одного типа нуклеиновой кислоты - ДНК или РНК. По этому признаку все вирусы разделены на два класса - ДНК-содержащие вирусы и РНК-содержащие вирусы.
Облигатный внутриклеточный паразитизм. Вирусы способны реплицироваться (размножаться) только внутри живых клеток, так как у них отсутствуют собственные системы, синтезирующие белок и генерирующие энергию.
Простое строение вириона (вирусной частицы). Вирион состоит из генома (ДНК или РНК), покрытого одной или двумя оболочками (капсидом и суперкапсидом). У вирусов отсутствуют такие клеточные элементы как цитоплазма, клеточные мембраны, рибосомы и др.
Дизъюнктивной (разобщенной) способ репродукции внутри клетки. При репродукции вирусов в разных частях инфицированной клетки синтезируются нуклеиновые кислоты и белки, которые затем объединяются в дочерние вирусные частицы. Синтез компонентов вирусных частиц происходит либо в цитоплазме, либо в цитоплазме и ядре клетки.
Морфологию и структуру вирусов изучают в основном с помощью электронной микроскопии. Препараты для электронной микроскопии готовят специальными методами:
методом напыления (на подложку из чистого углерода или коллодия наносят вируссодержащий материал, лиофильно высушивают и напыляют тяжелые металлы - в частности, палладий);
методом реплик (вируссодержащий материал заливают тонким слоем пластмассы и микроскопируют);
методом негативного контрастирования (вируссодержащий материал помещают на подложку, добавляют раствор уранилацетата, который попадает во все углубления вириона и создает для электронов непроницаемый фон).
Размеры вирионов определяют с помощью ультрафильтрации через фильтры с известным диаметром пор в нм или методом ультрацентрифугирования. Крупные вирионы можно увидеть в световом микроскопе в виде мелких внутриклеточных образований - включений (например, тельца Пашена при оспе, тельца БабешаНегри при бешенстве).
Вирусная частица называется вирионом. Выделяют следующие формы вирионов:
палочковидная форма (рисунок 1) характерна для вируса табачной мозаики;
пулевидная форма (рисунок 2) присуща вирусу бешенства;
сферическая форма (рисунок 3) отмечается у многих вирусов, в частности у герпесвирусов, вируса иммунодефицита человека;
нитевидная форма (рисунок 4) наблюдается у филовирусов;
овальная форма (рисунок 5) характерна для вируса натуральной оспы;
сперматозоидная форма (рисунок 6) отмечается у большинства вирусов бактерий (бактериофагов).


Рисунок 1 - Палочковидная форма вирионов. Электронная микроскопия.
Компьютерная окраска.
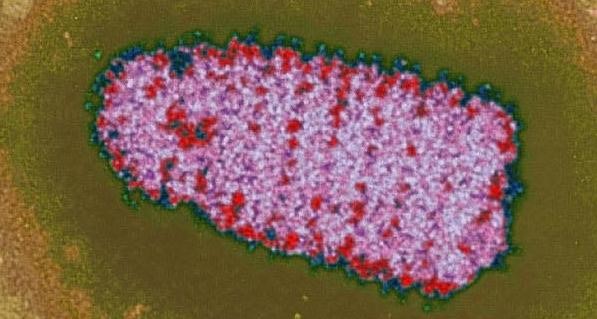

Рисунок 2 - Пулевидная форма вирионов. Электронная микроскопия.
Компьютерная окраска.

Рисунок 3 - Сферическая форма вирионов. Электронная микроскопия.

Рисунок 4 - Нитевидная форма вирионов. Электронная микроскопия. Компьютерная окраска.
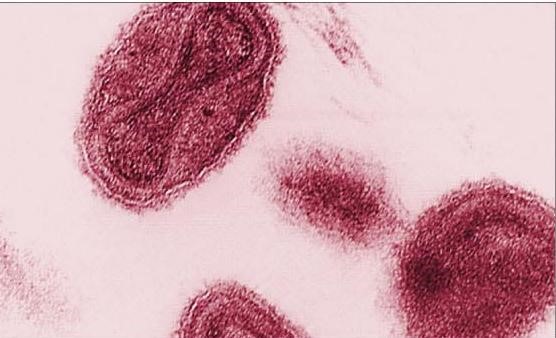

Рисунок 5 - Овальная форма вириона вируса натуральной оспы. Электронная микроскопия. Компьютерная окраска.
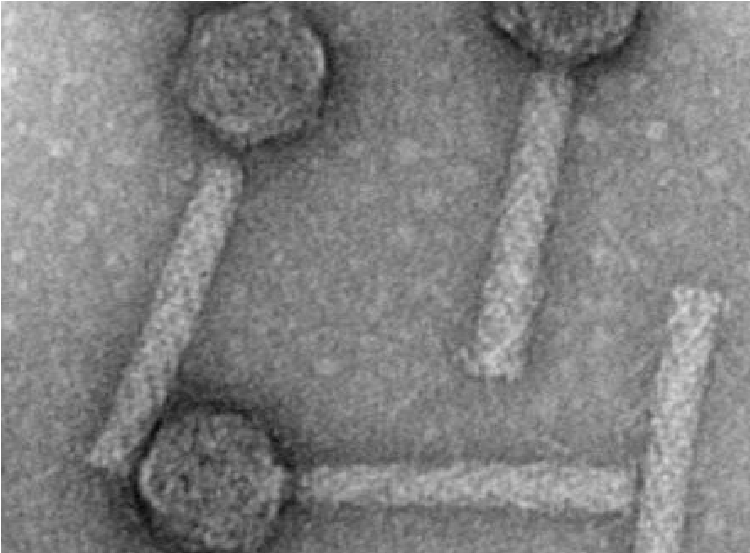
Рисунок 6 - Сперматозоидная форма вирионов. Электронная микроскопия бактериофагов.
Все вирусы существуют в двух качественно разных формах: внеклеточной - вирион и внутриклеточной - вирус. Таксономия этих представителей микромира основана на характеристике вирионов - конечной фазы развития вирусов.
Строение (морфология) вирусов.
1.Геном вирусов образуют нуклеиновые кислоты, представленные одноцепочечными молекулами РНК (у большинства РНК- вирусов) или двухцепочечными молекулами ДНК (у большинства ДНК- вирусов).
2.Капсид - белковая оболочка, в которую упакована геномная нуклеиновая кислота. Капсид состоит из идентичных белковых субъединиц - капсомеров. Существуют два способа упаковки капсомеров в капсид - спиральный (спиральные вирусы) и кубический (сферические вирусы).
При спиральной симметрии белковые субъединицы располагаются по спирали, а между ними, также по спирали, уложена геномная нуклеиновая кислота (нитевидные вирусы). При кубическом типе симметрии вирионы могут быть в виде многогранников, чаще всего - двадцатигранники - икосаэдры.
3.Просто устроенные вирусы имеют только нуклеокапсид, т.е. комплекс генома с капсидом и называются “голыми”.
4. У других вирусов поверх капсида есть дополнительная мембраноподобная оболочка, приобретаемая вирусом в момент выхода из клетки хозяина - суперкапсид. Такие вирусы называют “одетыми”.
Кроме вирусов, имеются еще более просто устроенные формы способных передаваться агентов - плазмиды, вироиды и прионы.
Строение вирусов
По строению различают два типа вирусных частиц – простые и сложные. В составе простых вирионов есть ДНК или РНК и белки. У сложных в суперкапсиде содержатся липиды, полисахариды.
Внутренняя структура простых и сложных вируосв сходна, сердцевина вируса – вирусный геном, который содержит от 3 до 100 и более генов.
Морфология и структура вирусов. Простые вирусы имеют одну белковую оболочку – капсид, который состоит из капсомеров – белковых молекул, форма укладки которых определяет тип симметрии. Капсид представлен -спиральными белками, способными к полимеризации.
Сложные вирусы имеют внешнюю оболочку – суперкапсид, расположенную поверх капсида. В состав суперкапсида входит внутренний белковый слой – М-белок, затем более объемный слой липидов и углеводов, извлеченных из клеточных мембран клетки хозяина. Вирусспецифические гликопротеиды проникают внутрь суперкапсида, образуя фигурные выпячивания (шипы, фибры), которые выполняют рецепторную функцию.
Различают 3 типа симметрии: 1) спиральный, когда капсомеры укладываются по спирали – винтообразная структура нуклеокапсида; 2) кубический (икосаэдрический), когда капсомеры укладываются по граням многогранника (12-20-гранника) – в основе лежит фигура икосаэдра (20-гранника). В зависимости от типа перегруппировки и числа субъединиц число капсомеров будет равным 30, 20, или 12. Вирионы со сложным капсидом, построенным более чем из 60 капсомеров, содержат группы из 5 субъединиц – пентамеры, или из 6 субъединиц – гексамеры; 3) смешанный тип симметрии (у бактериофагов).
Комплекс капсида и генома вируса называют нуклеокапсид. Сложные вирусы имеют суперкапсид (пеплос). Эта поверхностная оболочка вируса, состоит из липидов и белков клеточного происхождения.
Вирусные белки бывают: 1) структурные; 2) неструктурные.
Среди структурных различают: капсидные – входят в состав капсомеров и образуют футляр, защищающий нуклеиновую кислоту; суперкапсидные – это гликопротеиды, которые формируют шипы на поверхности суперкапсида и выполняют: адресную функцию – узнают чувствительную клетку и адсорбируются на ней; прикрепительныебелки, которые взаимодействуют со специфическими рецепторами клетки; белкислияния – способствуют слиянию вирусной и клеточной мембран и приводят к образованию симпластов; геномные – обладают антигенными свойсвами, участвуют во взаимодействии с клеткой.
Среди неструктурных белков различают: предшественники вирусных белков (нестабильные); РНК- и ДНК-полимеразы – участвуют в репликации вирусного генома; регуляторные белки – участвуют в репродукции вируса.
Функции белков:обладают антигенными и иммуногенными свойствами; участвуют в распознавании клетки и взаимодействии с ней; защищают геном от нуклеаз; обеспечивают тип симметрии.
Липиды входят в состав суперкапсида и представляют смесь нейтральных фосфо- и гликолипидов, многие из них – продукты мембраны клеток хозяина.
Они обусловливают инфекционность, чувствительность или устойчивость к эфиру; стабилизируют вирусную частицу.
Углеводы входят в состав гликопротеидов суперкапсида. Углеводы и липиды – составная часть гемагглютинина, который вызывает склеивание эритроцитов и обладает антигенной специфичностью.
Различают вирионные и вирусиндуцированные ферменты вирусов. К вирионным относят ферменты транскрипции и репликации (ДНК и РНК-полимеразы); обратную транскриптазу (у ретровирусов), АТФ-азы, эндо- и экзонуклеазы, нейраминидазы.
К вирусиндуцированным относятся ферменты, о которых имеется только информация в вирусном геноме, а появляются они в клетке. Это РНК-полимеразы тога-, орто-, пикорна- и парамиксовирусов; и ДНК-полимеразы у покс- и герпесвирусов.
Нуклеиновые кислоты обеспечивают наследственные признаки; являются хранителями генетической информации; необходимы для репродукции вирусов, многие из них могут вызывать инфекционный процесс самостоятельно, достаточно их проникновения в клетку.
Вирусная ДНК.Молекулярная масса равна 1,106-1,108 дальтон. ДНК может быть одно- или двунитчатой, фрагментированной и сверхспирализованной, линейной или кольцевой, содержит несколько сотен генов. В каждой нити ДНК есть нуклеотидные последовательности, а на концах есть прямые или инвертированные (повернутые на 180о) повторы, которые являются маркерами для отличия вирусной ДНК от клеточной. Эти повторы обеспечивают способность ДНК замыкаться в кольцо для последующих репликации, транскрибирования и встраивания в клеточный геном. Генетическая информация инфекционной ДНК транслируется на мРНК в клетке с помощью полимераз.
Вирусная РНКможет быть одно- и двунитчатой, линейной, кольцевой, фрагментированной. У РНК-содержащих вирусов генетическая информация закодирована в РНК таким же кодом, как в ДНК всех других вирусов и клеточных организмов. Вирусные РНК по своему химическому составу не отличаются от РНК клеточного происхождения, но характеризуются разной структурой.
Наряду с типичной для всех РНК однонитевой формой у ряда вирусов имеется двунитевая РНК. В составе однонитевых РНК имеются спиральные участки типа двойной спирали ДНК, образующиеся вследствие спаривания комлементарных азотистых оснований. Вирусы с однонитчатой РНК делятся на 2 группы: (+)РНК (положительный геном) и (-)РНК (отрицательный геном). Вирусная (+)РНК инфекционная и обладает функциями информационной РНК. Она может передовать генетическую информацию на рибосомы, как иРНК. Вирусы с отрицательным геномом не обладают инфекциозностью, т.к. нить (-)РНК выполняет только наследственную функцию и не обладает функцией иРНК. В зараженной клетке на матрице вирусной геномной РНК с помощью фермента транскриптазы осуществляется синтез РНК-комплементарной геному.
Нити (+)РНК вирусов в отличие от (-)РНК имеют специальные концы в виде «шапочки» для специфического узнавания рибосом.
Патогенность вирусов обусловлена совокупностью их свойств: способностью проникать в макроорганизм, связываться с клеточными мембранами и проникать в клетку, управлять метаболизмом и белоксинтезирующей функцией клетки, обеспечивать транскрипцию и репликацию собственного генома и осуществлять весь цикл репродукции вирусов. Все эти свойства зависят от генома вирусов и наличия соответствующих структурных белков и ферментов. Репродукция вирусов приводит к развитию патологии: цитопатогенному (разрушающему) действию, развитию воспаления, повреждению различных клеток и тканей.
Классификация вирусов
В силу своих особенностей вирусы выделены в отдельное надцарство Vira, в котором по типу нуклеиновой кислоты различают рибовирусы и дезоксирибовирусы (табл. 1).
Подцарства делятся на семейства, которые подразделяются на подсемейства и роды. Вид – совокупность вирусов, имеющих почти идентичные геном (ДНК или РНК), свойства и способность вызывать определенный патологический процесс. Названия семейства имеют окончание viridae, подсемейство – virinae, рода – virus.
Признаки, используемые для классификации вирусов: 1) тип нуклеиновой кислоты – ДНК или РНК; 2) их структура (однонитевая, двунитевая, линейная, кольцевая, фрагментированная, нефрагментированная с повторяющимися и инвертированными последовательностями); 3) структура, размеры, тип симметрии, число капсомеров; 4) наличие или отсутствие внешней оболочки (суперкапсида); 5) антигенная структура; 6) феномены генетических взаимодействий; 7) круг восприимчивых хозяев; 8) географическое распространение; 9) внутриядерная или цитоплазматическая локализация; 10) чувствительность к эфиру и детергентам; 11) путь передачи инфекции.
Для определения принадлежности к семейству ретровирусов обязательно учитывается наличие фермента обратной транскриптазы.
Вирусы, вызывающие инфекционные процессы у человека, входят в состав как ДНК-содержащих, так и РНК-содержащих вирусных семейств (см. табл. 1).
|
|
|
 Скачать 7.68 Mb.
Скачать 7.68 Mb.