Норм физ. 1 Строение плазматической мембраны, роль ионных каналов, молекул переносчиков, насосов, рецепторов. Виды транспорта веществ
 Скачать 95.91 Kb. Скачать 95.91 Kb.
|

Закон градиента, или аккомодацииЭтот закон гласит, что действие раздражителя зависит не только от абсолютной величины его силы, но и от скорости ее нарастания до порогового значения. Например, действие очень медленно нарастающего раздражения не вызывает возбуждения, так как раздражаемая ткань адаптируется к его влиянию. Этот факт объясняется тем, что при действии медленно возрастающего раздражителя в мембране возбудимой ткани увеличивается уровень деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения ответная реакция не возникает при любой силе раздражителя. Это явление получило название аккомодации. Причиной аккомодации является инактивация Na+-каналов, возникающая при медленной деполяризации клеточной мембраны, которая длится в течение 1 с и более. Способность к аккомодации у различных структур неодинакова. Она высокая у нервных волокон и низкая у сердечной мышцы, гладких мышц кишечника и желудка. Закон силы-времени, раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он действует. Чем больше ток, тем меньше времени он должен действовать на возбудимые ткани, чтобы вызвать возбуждение. Существует зависимость между силой раздражения и временем его действия. Эта зависимость выражается в виде кривой силы и времени. Кривая показывает, что каким бы сильным ни был бы раздражитель, он должен действовать определенный период времени. Если временной отрезок маленький, то ответная реакция не возникает. Если раздражитель слабый, то бы как длительно он ни действовал, ответная реакция не возникает. Сила раздражителя постепенно увеличивается, и в определенный момент возникает ответная реакция ткани. Эта сила достигает пороговой величины и называется реобазой (минимальной силой раздражения, которая вызывает первичную ответную реакцию). Время, в течение которого действует ток, равный реобазе, называется полезным временем. 6) Классификация нервных волокон Классификация нервных волокон Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в таблице 4.1. Нервные волокна всех групп обладают общими свойствами: нервные волокна практически неутомляемы; нервные волокна обладают высокой лабильностью, т.е. могут воспроизводить потенциал действия с очень высокой частотой. Существует три закона проведения раздражения по нервному волокну. Закон анатомо-физиологической целостности. Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно. Закон изолированного проведения возбуждения. Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах. В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки. В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна. Закон двустороннего проведения возбуждения. Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно. В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении. Так, при распространении возбуждения по безмиелиновому нервному волокну местные токи, которые возникают между его возбужденным и невозбужденным участками, вызывают деполяризацию мембраны и генерацию потенциала действия. Затем локальные токи возникают уже между возбужденным участком мембраны и ближайшим невозбужденным участком. Многократное повторение этого процесса способствует распространению возбуждения вдоль нервного волокна. Так как в процесс возбуждения последовательно вовлекаются все участки мембраны волокна, то такой механизм проведения возбуждения называется непрерывным. Непрерывное проведение потенциала действия происходит в мышечных волокнах и в безмиелиновых нервных волокнах типа С. Наличие у миелиновых нервных волокон участков без этой миелиновой оболочки (перехваты Ранвье), обусловливает специфический тип проведения возбуждения. В этих волокнах местные электрические токи возникают между соседними перехватами Ранвье, разделенными участком волокна с миелиновой оболочкой. И возбуждение «перепрыгивает» через участки, покрытые миелиновой оболочкой, от одного перехвата к другому. Такой механизм распространения возбуждения называется сальтаторным (скачкообразным), или прерывистым. Скорость сальтаторного проведения возбуждения гораздо выше, чем в безмиелиновых волокнах, так как в процесс возбуждения вовлекается не вся мембрана, а только ее небольшие участки в области перехватов. «Перепрыгивание» потенциала действия через миелиновый участок возможно потому, что его амплитуда в 5-6 раз превышает величину, необходимую для возбуждения соседнего перехвата Ранвье. Иногда потенциал действия способен «перепрыгивать» даже через несколько межперехватных промежутков. 7) Нервно – мышечный синапс Нервно-мышечный синапс образован окончанием аксона моторного нейрона и мышечным волокном поперечно-полосатой мускулатуры. Структурами, непосредственно образующими синапс, являются пресинаптическая мембрана аксонной терминали, синаптическая щель и постсинантическая мембрана, являющаяся частью плазматической мембраны (сарколеммы) постсинаптической мышечной клетки. Пресинаптической мембраной называют часть не покрытой миелином мембраны аксонной терминали, обращенной в синаптическую щель. Пресинаптическая терминаль формируется тонкой веточкой аксона, которая, подходя к мышечному волокну, образует утолщение (пуговку, бляшку, бутон). В пресинаптической терминали находятся синаптические пузырьки (везикулы), содержащие запасы медиатора ацетилхолина. Везикул может быть до нескольких тысяч. Их диаметр около 40 им, и в каждой содержится несколько тысяч молекул медиатора. В отсутствие поступления нервных импульсов везикулы с помощью белка синапсина связаны с цитоскелетом и малоподвижны. В пресинаптической терминали имеются также митохондрии, обеспечивающие выработку АТФ, ацетил СоА, белки цитоскелета, микротрубочки и микронити, по которым в терминаль из тела нейрона перемещается фермент ацетилхолинтрансфсраза. При участии этого фермента из ацетил СоА и холина образуется ацетилхолин. 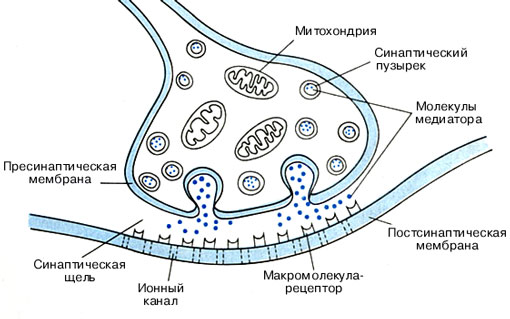 Потенциал концевой пластинки В нервно-мышечном синапсе ( рис. 382.1 ) ацетилхолин синтезируется в окончаниях двигательных нервов и накапливается в пузырьках. Когда в окончание приходит потенциал действия, ацетилхолин из 150-200 пузырьков высвобождается в синаптическую щель и связывается с холинорецепторами (холинорецепторы нервно-мышечных синапсов принадлежат к N-холинорецепторам ), плотность которых особенно высока на гребнях складок постсинаптической мембраны. Каналы, сопряженные с холинорецепторами, открываются, в клетку входят катионы (в основном Na+), и происходит деполяризация постсинаптической мембраны, называемая потенциалом концевой пластинки. Поскольку этот потенциал в норме всегда сверхпороговый, он вызывает потенциал действия, распространяющийся по мышечному волокну и вызывающий сокращение. Потенциал концевой пластинки короткий, так как ацетилхолин, во-первых, быстро отсоединяется от рецепторов, во-вторых - гидролизуется АХЭ . 8)режимы и типы сокращений скелетных мышц характеристика двигательных единиц физиологические особенности гладких мышц Режимы и виды мышечных сокращений. Изотонический режим — мышца укорачивается при отсутствии возрастания ее напряжения. Такое сокращение возможно только для изолированной (удаленной из организма) мышцы. Изометрический режим — напряжение мышцы возрастает, а длина практически не уменьшается. Такое сокращение наблюдается при попытке поднять непосильный груз. Ауксотонический режим мышца укорачивается и возрастает ее напряжение. Такое сокращение чаще всего наблюдается при осуществлении трудовой деятельности человека. Вместо термина "ауксотонический режим" часто применяется названиеконцентрический режим. Выделяют два вида мышечных сокращений: одиночное и те- таническое. Одиночное мышечное сокращение проявляется в результате развития одиночной волны возбуждения в мышечных волокнах. Этого можно достичь при воздействии на мышцу очень коротким (около 1 мс) стимулом. В развитии одиночного мышечного сокращения выделяют латентный период, фазу укорочения и фазу расслабления. Сокращение мышцы начинает проявляться через 10 мс от начала воздействия раздражителя. Этот временной интервал называют латентным периодом (рис.5.1). Затем последует развитие укорочения (длительность около 50 мс) и расслабления (50—60 мс). Считается, что на весь цикл одиночного мышечного сокращения затрачивается в среднем 0,1 с. Но следует иметь в виду, что длительность одиночного сокращения у разных мышц может сильно варьировать. Она также зависит от функционального состояния мышцы. Скорость сокращения и особенно расслабления замедляется при развитии утомления мышцы. К быстрым мышцам, имеющим короткий период одиночного сокращения, относятся мышцы языка и смыкающие веко.  Рис. 5.1. Временные соотношения разных проявлений возбуждения волокна скелетной мышцы: а — соотношение потенциала действия, выхода Са2+в саркоплазму и сокращения: / — латентный период; 2 —укорочение; 3 — расслабление; б - соотношение потенциала действия, сокращения и уровня возбудимости Под влиянием одиночного раздражителя вначале возникает потенциал действия и лишь затем начинает развиваться период укорочения. Оно продолжается и после окончания реполяризации. Восстановление исходной поляризации сарколеммы свидетельствует и о восстановлении возбудимости. Следовательно, на фоне развивающегося сокращения в мышечных волокнах можно вызвать новые волны возбуждения, сократительный эффект от которых будет суммироваться. Тетаническим сокращением или тетанусом называют сокращение мышцы, появляющееся в результате возникновения в моторных единицах многочисленных волн возбуждения, сократительный эффект от которых суммируется по амплитуде и времени. Различают зубчатый и гладкий тетанус. Для получения зубчатого тетануса надо стимулировать мышцу с такой частотой, чтобы каждое последующее воздействие наносилось после фазы укорочения, но до момента окончания расслабления. Гладкий тетанус получается при более частых раздражениях, когда последующие воздействия наносятся во время развития укорочения мышцы. Например, если фаза укорочения у мышцы составляет 50 мс, а фаза расслабления — 60 мс, то для получения зубчатого тетануса необходимо раздражать эту мышцу с частотой 9—19 Гц, для получения гладкого — с частотой не менее 20 Гц. Несмотря  Характеристика двигальных единиц. Двигательная единица представляет собой систему из мотонейрона и иннервируемых им мышечных волокон ( 21). Двигательная единица работает как одно целое. Все мышечные волокна, входящие в состав данной двигательной единицы, сокращаются практически одновременно. Мотонейроны бывают большие и малые. Малые мотонейроны имеют тонкие аксоны и иннервируют небольшое количество (десятки) мышечных волокон, образуя малые двигательные единицы. Большие мотонейроны имеют толстые аксоны, которые иннервируют большое количество мышечных волокон (до нескольких тысяч), образуя большие двигательные единицы. Малые двигательные единицы входят в состав главным образом мелких мышц (пальцев рук, лица и др.), однако они входят также и в состав крупных мышц. Малые двигательные единицы обеспечивают быстрые и тонкие движения (например, движения пальцев рук). Большие двигательные единицы входят в состав преимущественно крупных мышц туловища и конечностей. Эти мышцы осуществляют относительно менее тонкие и более медленные движения, чем, например, движения пальцев рук. Малые мотонейроны (низкопороговые) возбуждаются легче и быстрее по сравнению с большими (высокопороговыми). Большинство скелетных мышц человека состоит как из медленных, так и из быстрых мышечных волокон. Поэтому волокна одной и той же мышцы могут выполнять относительно и более быстрые, и менее быстрые движения. Это зависит от того, какие двигательные единицы (быстрые или медленные) будут включены в работу. Мышцы бывают быстрые и медленные. Так, например, внутренняя прямая мышца глаза относится к быстрым, а камбаловидная к медленным мышцам. Длительность волны сокращения первой равна 7,5 мс, а второй 75 мс, т. е. в 10 раз больше. 9) Механизм сокращения скелетных мышц. Теория скольжения: роль ионов кальция, регуляторных и сократительных белков в мышечном сокращении и расслаблении Изменение механического состояния миофибриллярного сократительного аппарата мышечных волокон называется сокращением. При произвольной внутренней команде сокращение мышцы человека начинается примерно через 0.05 с. За это время моторная команда передается от коры больших полушарий к мотонейронам спинного мозга и по двигательным волокнам к мышце. Подойдя к мышце, процесс возбуждения должен с помощью медиатора преодолеть нервно-мышечный синапс, что занимает примерно 0.5 мс. Выделившийся в синаптическую щель медиатор прикрепляется к рецепторам постсинаптической мембраны и вызывает в ней явления деполяризации. Расслабление мышечного волокна связано с работой особого механизма - "кальциевого насоса", который обеспечивает откачку ионов Са2+ из миофибрилл обратно в трубочки саркоплазматического ретикулума. По теории скольжения, в основе сокращения лежит механическое взаимодействие между миозиновыми и актиновыми миофиламентами благодаря образованию между ними в период активности поперечных мостиков. Продвижение тонких миофиламентов относительно толстых сопровождается приближением поперечных мостиков к зоне расположения АТФ-азы, что вызывает гидролиз АТФ и соответственно разрушение мостиков. Данные о роли ионов кальция в сократительной активности мышц накапливались довольно медленно. Кальций активен в саркоплазме при такой низкой концентрации. Количественная зависимость между концентрацией свободного Са2+ в саркоплазме и силой мышечного сокращения была установлена сравнительно недавно. Саркоплазматический ретикулум - это внутриклеточная мембранная система взаимосвязанных уплощенных пузырьков и канальцев (цистерн), которая окружает саркомеры. На внутренней мембране расположены белки, способные связывать ионы кальция. Основная функция СР заключается в регуляции содержания ионов кальция в пространстве между актином и миозином. Энергетика мышечного сокращения.Сокращение мышцы - это движения мостиков, которым требуется энергия. Запас энергии в мышце ограничен, поэтому при работе мышцы требуется постоянное ее восполнение. Основным источником энергии в организме служат молекулы АТФ, с помощью них происходят все энергетические процессы в клетке. При взаимодействии с нитью актина, головки миозиновых мостиков расщепляют молекулу АТФ, получая тем самым энергию для сокращения. Но запаса молекул АТФ в цитоплазме мышечной клетки хватает лишь на непродолжительную мышечную работу. Формы сокращения мышц (изотоническая, изометрическая, смешанная). Особенности одиночных и тетанических сокращений медленных и быстрых мышечных волокон. Связь исходной длины и силы сокращения скелетной мышцы. Зависимость между и скоростью сокращения мышц. · Динамическая форма: 1)Изотонический тип - (мышца укорачивается, но не изменяет своего напряжения). Например, ходьба. 2) Эксцентрический тип. Если нагрузка на мышцу больше, чем ее напряжение, то мышца растягивается. Например, при опускании тяжелого предмета. · Статическая форма мышечного сокращения. Эта форма наблюдается при поддержании позы или преодолении силы земного притяжения. · Форма смешанная: Деление на формы и типы мышечных сокращений является условным т.к. все сокращения являются смешанными. При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается одиночным сокращением. Сокращение целой мышцы зависит от формы сокращения отдельных ДЕ и их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно поддерживая общее напряжение мышцы на заданном уровне. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения. В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды. |