1. Строение прокариотической клетки
 Скачать 198.54 Kb. Скачать 198.54 Kb.
|

1. Строение прокариотической клеткиК прокариотам относятся архебактерии, бактерии и синезеленые водоросли. Прокариоты — одноклеточные организмы, у которых отсутствуют структурно оформленное ядро, мембранные органоиды и митоз. Размеры клеток не более 10 мкм, обычно 0,5-3 мкм, отсутствует клеточный центр, отсутствует большинство органелл, отсутствующие органеллы заменяет выросты цитоплазматической мембраны- мезосомы, отсутствует циклоз – движение цитоплазмы, рибосомы прокариотической клетки существенно отличаются от рибосом эукариот, отсутствует ядро (есть кольцевая молекула ДНК единственной хромосомы, лишенная белков-гистонов). К прокариотам относят архебактерии (наиболее древние), истинные бактерии и сине-зеленые водоросли. Среди прокариотов есть аэробы, анаэробы, автотрофы и гетеротрофы. Прокариоты определяют границы жизни на Земле, обеспечивают круговорот многих веществ в природе. 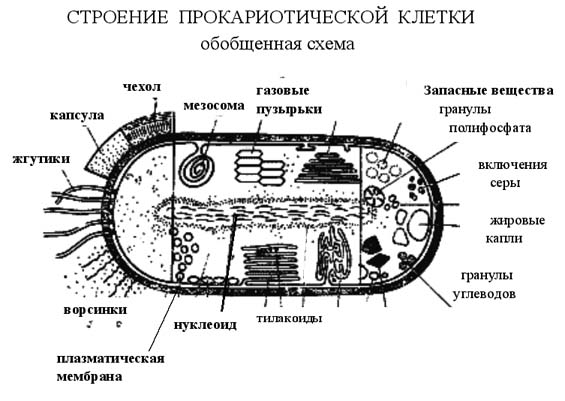 2.Общий план строения эукариотической животной клетки Эукариотические клетки имеют структурно оформленное ядро, возникли на базе прокариотических клеток благодаря эндосимбиозу разных прокариотических клеток. Размеры эукариотических клеток тканей животных и растений варьируют от 10 до 100 мкм. Основные компоненты – оболочка, цитоплазма, морфологически оформленное ядро. Генетический материал сосредоточен преимущественно в хромосомах ядра. 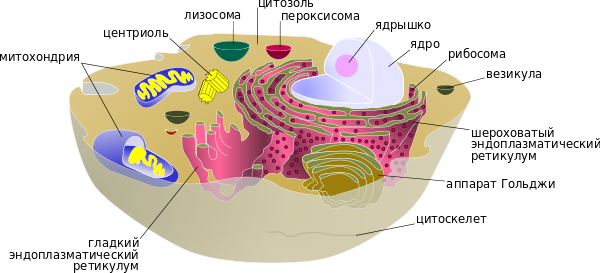 3. Системы жизнеобеспечения клетки САМОСОХРАНЕНИЕ (система мембран), САМОРЕГУЛЯЦИЯ (система получения и превращения энергии), САМОВОСПРОИЗВЕДЕНИЕ (репликация, транскрипция и трансляция), Система мембран (цитоплазматическая мембрана, мембранные органеллы). Система авторепродукции (воспроизведение себе подобных) включает ДНК, РНК, рибосомы, множество ферментов. Система получения и трансформации энергии: митохондрии и хлоропласты. 4. Строение и функции цитоплазматической мембраны 1. Фосфолипидный бислой (заряженные «головки» – снаружи, незаряженные хвосты – внутри). 2. Белки (ферменты, рецепторы, переносчики и др.) встроены в фосфолипидный остов. 3. Снаружи к мембране примыкают липиды (липопротеиды) и углеводы (гликопротеиды), изнутри – белки. Гликокаликс образован комплексами полисахаридов с белками – гликопретеинами и жирами – гликолипидами. Процентное содержание компонентов мембраны: белков в мембране колеблется от 40 до 70%, липидов – от 25 до 60%, углеводов – от 5 до 10%. Функции цитоплазматической мембраны: защитная, регуляция проникновения веществ в клетку (К+/Na+-насос), рецепторная функция – восприятие сигналов, антигенная функция (гликопротеиды мембран являются антигенами (эритроцитарные антигены – группы крови), электрогенная. Компартментация – это разделения на ячейки, отличные деталями химического состава. 5. Транспорт различных веществ через цитоплазматическую мембрану. Механизмы транспорта. Существует несколько механизмов транспорта веществ через мембрану: Диффузия – проникновение веществ через мембрану по градиенту концентрации (из области высшей концентрации в область низшей). Диффузный транспорт веществ осуществляется при участии белков мембраны, в которых имеются молекулярные поры (вода, ионы), либо при участии липидной фазы (для жирорастворимых веществ). Облегченная диффузия– специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом или молекулой и переносят их через мембрану. Активный транспорт– этот механизм сопряжен с затратами энергии и служит для переноса веществ против их градиента концентрации. Он осуществляется специальными белками-переносчиками, образующими так называемые ионные насосы. Наиболее изученным является Na+/К+ - насос в клетках животных, активно выкачивающий ионы Na наружу, поглощая при этом ионы К+. Пассивный транспорт: Эндоцитоз- транспорт макромолекул, их комплексов и частиц внутрь клетки. При эндоцитозе определенный участок плазмалеммы захватывает внеклеточный материал, заключая его в мембранную вакуоль, возникшую за счет выпячивания мембраны. В дальнейшем такая вакуоль соединяется с лизосомой, ферменты которой расщепляют макромолекулы до мономеров. Эндоцитоз разделяют на фагоцитоз (захват и поглощение твердых частиц) и пиноцитоз (поглощение жидкости). Путем эндоцитоза осуществляется питание гетеротрофных протистов, защитные реакции организмов (лейкоциты поглощают чужеродные частицы) и др. Экзоцитоз– (экзо - наружу) благодаря ему клетка выводит внутриклеточные продукты или непереваренные остатки, заключенные в вакуоли или пузырьки. Пузырек подходит к цитоплазматической мембране, сливается с ней, а его содержимое выделяется в окружающую среду. Так выделяются пищеварительные ферменты, гормоны, гемицеллюлоза и др. 6. Строение ядра клетки и функция его основных органоидов Основные компоненты ядра: ядерная оболочка, кариоплазма или ядерный сок, хромосомы – хроматин, ядрышки. Ядерная оболочка состоит из двух мембран (наружной и внутренней), между которыми находится перинуклеалярное пространство. Наружная мембрана соединена с каналами ЭПС. Обе мембраны пронизаны порами. Через поры осуществляется транспорт веществ из ядра в цитоплазму: субчастицы рибосом, иРНК и др. Ядерная оболочка разобщает транскрипцию, протекающую в ядре, от трансляции белка в цитоплазме. Кариоплазма содержит ферменты и нуклеотиды, необходимые для синтеза нуклеиновых кислот и субчастиц рибосом. В ней располагаются хроматин и ядрышки (одно или несколько) и происходит процессинг (созревание) иРНК, тРНК и рРНК. Хромосомы представляют собой комплекс, состоящий из ДНК (40%) и белков- гистонов (60%), богатых аминокислотами аргинином и лизином. Различают эухроматин (максимально деконденсированные или рыхлые участки хромосом) и гетерохроматин (всегда конденсированные участки хромосом, которые реплицируются позже, чем эухроматин). Ядрышки непостоянные структуры: они демонтируются в начале деления клетки и вновь появляются к концу его. Химический состав: рРНК, белки и ДНК. Образуются на определенных участках хромосом – ядрышковых организаторах. В кариотипе человека ядрышкообразующие хромосомы относятся к группе D: 13, 14, 15 пары и к группе G: 21, 22 пары. В ядрышке происходит синтез рРНК и образование субчастиц рибосом. 7. Строение и функции ДНК ДНК состоит из нуклеотидов, в состав которых входят сахар – дезоксирибоза, фосфат и одно из азотистых оснований – пурин (аденин или гуанин) либо пиримидин (тимин или цитозин). Молекулы ДНК включают в себя 2 полинуклеотидные цепи, соединенные друг с другом азотистыми основаниями с помощью водородных связей по принципу комплементарности (аденин -2вод.связи-тимин, гуанин-3вод.связи-цитозин). Цепи антипараллельны: 5’-конец одной цепи соединяется с 3’-концом другой цепи. Чаще всего спирали правозакрученные. В структурной организации молекулы ДНК можно выделить первичную структуру – полинуклеотидную цепь, вторичную структуру – две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями, и третичную структуру – трехмерную спираль с приведенными выше пространсвенными характеристиками. Функции ДНК – сохранение и передача наследственной информации от клетки к клетке, от организма к организму (в основе этой функции лежит репликация ДНК); регуляция всех процессов, протекающих в клетке (в основе этой функции лежит транскрипция). Свойства: способность к самокопированию (репликации), к молекулярному восстановлению (репарации). Функции ДНК: * Молекулы ДНК хранят (содержат) наследственную информацию (программу) о структуре специфических для каждого организма белков. * Молекулы ДНК обеспечивают передачу наследственной информации от клетки к клетке, от организма к организму. * Молекулы ДНК участвуют в реализации генетической информации, т. е. участвуют в процессе синтеза полипептидов. 8. Репликация ДНК. Ферменты репликации Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа:
ДНК – полимераза ДНК-полимераза — фермент, участвующий в репликации ДНК. Ферменты этого класса катализируют полимеризацию дезоксирибонуклеотидов вдоль цепочки нуклеотидов ДНК, которую фермент «читает» и использует в качестве шаблона. Тип нового нуклеотида определяется по принципу комплементарности с шаблоном, с которого ведётся считывание. Собираемая молекула комплементарна шаблонной моноспирали и идентична второму компоненту двойной спирали. Выделяют ДНК-зависимую ДНК-полимеразу, использующую в качестве матрицы одну из цепей ДНК, и РНК-зависимую ДНК-полимеразу, способную также к считыванию информации с РНК (обратная транскрипция). ДНК-полимеразу считают холоферментом, поскольку для нормального функционирования она требует присутствия ионов магния в качестве кофактора. В отсутствии ионов магния о ней можно говорить как об апоферментe. ДНК-полимераза начинает репликацию ДНК, связываясь с отрезком цепи нуклеотидов. Среднее количество нуклеотидов, присоединяемое ферментов ДНК-полимеразой за один акт связывания/диссоциации с матрицей, называют процессивностью. ДНК – лигазы Лигаза — фермент, катализирующий соединение двух молекул с образованием новой химической связи (лигирование). При этом обычно происходит отщепление (гидролиз) небольшой химической группы от одной из молекул. Лигазы относятся к классу ферментов EC 6. В молекулярной биологии лигазы разделяют на две большие группы — РНК-лигазы и ДНК-лигазы. ДНК-лигаза, осуществляющая репарацию ДНК ДНК-лигазы — ферменты, катализирующие ковалентное сшивание цепей ДНК в дуплексе при репликации, репарации и рекомбинации. Они образуют фосфодиэфирные мостики между 5'-фосфорильной и 3'-гидроксильной группами соседних дезоксинуклеотидов в местах разрыва ДНК или между двумя молекулами ДНК. Для образования этих мостиков лигазы используют энергию гидролиза пирофосфорильной связи АТФ. Один из самых распространённых коммерчески доступных ферментов — ДНК-лигаза бактериофага Т4. ДНК – геликазы ДНК геликазы - ферменты раскручивающие двуцепочечную спираль ДНК с затратой энергии гидролиза трифосфатов NTP. Образуемая одноцепочечная ДНК участвует в различных процессах, таких как репликация, рекомбинация, и репарация. ДНК геликазы необходимы для репликации, репарации, рекомбинации и транскрипции. Геликазы присутствуют во всех организмах. ДНК-топоизомеразы ДНК-топоизомеразы—ферменты, изменяющие степень сверхспиральности и тип сверхспирали. Путём одноцепочечного разрыва они создают шарнир, вокруг которого нереплецированный дуплекс ДНК, находящейся перед вилкой, может свободно вращаться. Это снимает механическое напряжение, возникающее при раскручивании двух цепей в репликативной вилке, что является необходимым условием для её непрерывного движения. Кроме того, топоизомеразы (типа II) обеспечивают разделение или образование катенанов - сцепленных кольцевых ДНК (образуются в результате репликации кольцевой ДНК), а также устранение узлов и спутанных клубков из длинной линейной ДНК. Существует два типа топоизомераз. Топоизомеразы типа I уменьшают число сверхвитков в ДНК на единицу за один акт. Эти топоизомеразы надрезают одну из двух цепей, в результате чего фланкирующие дуплексные области могут повернутся вокруг интактной цепи, и затем воссоединяют концы разрезанной цепи. Эта реакция не требует энергии АТФ, т.к. энергия фосфодиэфирной связи сохраняется благодаря тому, что тирозиновый остаток в молекуле фермента выступает то в роли акцептора, то в роли донора фосфорильного конца разрезанной цепи. Топоизомеразы типа II вносят временные разрывы в обе комплиментарные цепи, пропускают двухцепочечный сегмент той же самой или другой молекулы ДНК через разрыв, а затем соединяют разорванные концы. В результате за один акт снимаются два положительных или отрицательных сверхвитка. Топоизомеразы типа II тоже используют тирозиновые остатки для связывания 5¢-конца каждой разорванной цепи в то время . когда другой дуплекс проходит через место разрыва. Праймаза Праймаза—фермент, обладающий РНК-полимеразной активностью; служит для образования РНК-праймеров, необходимых для инициации синтеза ДНК в точке ori и дальнейшем для синтеза отстающей цепи. 9. Строение и функции РНК РНК состоит из нуклеотидов, в состав которых входят сахар – рибоза, фосфат и одно из азотистых оснований (аденин, урацил, гуанин, цитозин). Образует первичную, вторичную и третичную структуры аналогично таким же у ДНК. Информация о последовательности аминокислот белка содержится в информационных РНК (иРНК, мРНК). Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК. Рибосомальные РНК (рРНК)— каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки. Функции: способность к самовоспроизведению, способность сохранять свою организацию постоянной, способность приобретать изменения и воспроизводить их. 10. Структура и свойства генетического кода Генетический код –Определенный набор и порядок расположения аминокислот в пептидных цепях. В многообразии белков, существующих в природе, было обнаружено около 20 различных аминокислот. Для их шифровки достаточное количество сочетаний нуклеотидов может обеспечить лишь триплетный код,в котором каждая аминокислота шифруется тремя стоящими рядом нуклеотидами, из четырех нуклеотидов образуется 43 = 64 триплета. Из 64 возможных триплетов ДНК 61 кодирует различные аминокислоты; оставшиеся 3 получили название бессмысленных, или «нонсенс-триплетов». Они не шифруют аминокислот и выполняют функцию знаков препинания при считывании наследственной информации. К ним относятся АТТ, АЦТ, АТЦ. Свойства генетического кода:вырожденность -явная избыточность кода, многие аминокислоты шифруются несколькими триплетами. Это свойство имеет очень важное значение, так как возникновение в структуре молекулы ДНК изменений по типу замены одного нуклеотида в полинуклеотидной цепи может не изменить смысла триплета. Возникшее новое сочетание из трех нуклеотидов кодирует ту же самую аминокислоту. Специфичность -каждый триплет способен кодировать только одну определенную аминокислоту. Универсальность - полное соответствие кода у различных видов живых организмов свидетельствует о единстве происхождения всего многообразия живых форм на Земле в процессе биологической эволюции. Непрерывность и неперекрываемость кодонов при считывании–последовательность нуклеотидов считывается триплет за триплетом без пропусков, при этом соседние триплеты не перекрывают друг друга, т.е. каждый отдельный нуклеотид входит в состав только одного триплета при заданной рамке считывания. Доказательством неперекрываемости генетического кода является замена только одной аминокислоты в пептиде при замене одного нуклеотида в ДНК . 11. Особенности структурной организации гена эукариот Структура гена эукариот:имеют мозаичную структуру: состоят из кодирующих (экзонов) и некодирующих (интронов) участков. Количество интронов – 1–60; количество нуклеотидов в них – десятки, тысячи и более. Например, ген фенилаланингидрооксилазы – 13 экзонов и 12 интронов (90 тыс.н.п.). Преимущества мозаичной структуры гена эукариот: повышается их информационную емкость (один ген может кодировать несколько полипептидов), увеличивается степень комбинативной изменчивости, обеспечивается более совершенная регуляция функции генов. Интроны регулируют процессинг иРНК. Функциональная единица генома эукариот включает один структурный ген (транскрибируемая зона) и множество регуляторных участков ДНК (промотор с ТАТА-блоком, энхансер, сайленсер, трейлер с кодонами – терминаторами и др.). Экзоны – нуклеотидные последовательности, кодирующие аминокислоты. Интроны – не кодирующие нуклеотидные последовательности (их от 2 до 7 на ген). Промотор (Р) - сайт для соединения с РНК-полимеразой. Сайленсер – ослабляет транскрипцию. Энхансер – усиливает транскрипцию. Зона кэпирования (К) – для формирования в зрелой иРНК КЭПа - метиловой «шапочки». Зона полиаденилирования (А) – для формирования в зрелой иРНК полиаденилового «хвоста». Зона терминации транскрипции (Т). 12.Свойство гена 1. Ген дискретен в своем действии, т. е. обособлен в своей активности от других генов. 2. Ген специфичен в своем проявлении, т. е. отвечает за строго определенный признак или свойство организма. 3. Ген может действовать градуально, т. е. усиливать степень проявления признака при увеличении числа доминантных аллелей (дозы гена ). 4. Один ген может влиять на развитие разных признаков — это множественное, или плейотропное, действие гена . 5. Разные гены могут оказывать одинаковое действие на развитие одного и того же признака (часто количественных признаков ) — это множественные гены, или полигены. 6. Ген может взаимодействовать с другими генами, что приводит к появлению новых признаков . Такое взаимодействие осуществляется опосредованно — через синтезированные под их контролем продукты своих реакций. 7. Действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или воздействием различных факторов внешней среды. 13-15. Этапы экспрессии гена эукариот: претранскрипционный, транскрипция, процессинг-сплайсинг, транспорт иРНК через ядерную мембрану, трансляция, посттрансляционный ДНК - про-иРНК - иРНК - белок Этапы экспрессии генов
Претранскрипционный этап - активация генов. Активаторы: рН, ионы, БАВ, метаболиты и др. Регуляторы активности генов в претранскрипционный период: внутриклеточные (pH ±, Ионы ±,Белки (гистоновые, негистоновые) ±,Метаболиты ±, Медиаторы ±, БАВ ±, «Прыгающие гены» (их локализация)). Внеклеточные (Медиаторы ±, Гормоны ±,Др. раздражители). Все вызывают изменения МП плазм. мембр. ядерная МБР хромосомы. Функции регуляторов генной активности (в зоне действия): освобождают ДНК от белков, деконденсация ДНП, Ослабляют водородные связи, активируют РНК-полимеразу, блокируют белок-репрессор – освобождают оператор от блока (у прокариот), активируют регуляторные зоны. Транскрипция– синтез РНК на матрице ДНК, начинается с присоединения РНК-полимеразы к «своему» промотору (ТАТА-блоку); траскрипция происходит с одной полипептидной цепи ДНК (кодогенной), на которой находится промотор. Направление транскрипции 3’→5’. Нуклеотидная цепь иРНК растет в направлении 5’→3’.Транскрибируются все экзоны и интроны (включая зону копирования, кодон-инициатор, зоны терминации и полиаденилирования) структурного гена. В итоге образуется про-иРНК, содержащая кодирующие и некодирующие нуклеотидные последовательности. Фермент – РНК-полимераза. Процессинг про-иРНК:Кэпирование – образование КЭП на 5’-конце про-иРНК(служит для присоединения к рибосоме). Полиаденилирование на 3’-конце (присоединение поли-А или поли-У) - придает устойчивость иРНК. Процессинг – сплайсинг – «вырезание» интронов и «сшивание» (ферментами-рибозимами) экзонов. Транспорт иРНК через ядерную мембрану- транспорт зрелой иРНК через ядерную мембрану. Трансляция– синтез белков рибосомами на матрице иРНК. Зрелая иРНК 5’-концом (КЭП) подходит к полисомам и протягивается через них, транслируются кодон-инициатор и все нуклеотиды кодирующей зоны, кодоны-терминаторы не транслируются. Посттрансляционный этап - образование функционально активного белка: у эукариот «отрезается» метионин или триптофан, формируется вторичная, третичная, а для многих белков и четвертичная структура, присоединяются др. группировки и т.д. 16. Химическая организация хромосом. Уровни компактизации ДНП: нуклеосомный, фибрилла, интерфазная хромосома, метафазная хромосома Химический состав хромосом:ДНК и белки, которые образуют нуклеопротеиновый комплекс—хроматин. Белки (гистоны) составляют значительную часть вещества хромосом. На их долю приходится около 65% массы этих структур. Помимо ДНК и белков в составе хромосом обнаруживаются также РНК, липиды, полисахариды, ионы металлов. Нуклеосомный: 4 класса гистоновых белков (из групп Н3, Н4, Н2А, Н2В) формируют ядро-нуклеус, вокруг которого ДНК делает 1.5 оборота – около 140 нуклеотидных пар, между нуклеосомами 50–70 нуклеотидных пар). За счет нуклеосомной организации ДНК укорачивается 6–7 раз. Фибриллярный: при образовании фибриллы нуклеосомы сближаются за счет разности зарядов «хвоста» гистона I (HI) и фосфатной группы ДНК. Гистоновый белок Н1 связывается с линкерной ДНК. Интерфазные хромосомы: максимально деконденсированные, 1 молекула ДНК, функционально активны, не видны в световой микроскоп, видны глыбки хроматина: у жен.- тельца Барра, у мужчин: У-хромосом. Метафазные хромосомы: максимально конденсированные, 2 хроматиды, функционально неактивны, видны в световой микроскоп. 17. Строение и функции метафазных хромосом Первичная перетяжка – центромера (С) - Локализована во всех хромосомах человека. В области центромеры расположен гетерохроматин. Центромера делит хромосому на два плеча: короткое «р» и длинное «q». Функции: во время митоза к центромере прикрепляются нити веретена деления, растаскивающие хромосомы к полюсам клетки. Вторичная перетяжка (h)- Вторичная перетяжка р-плеча (ph) представлена ядрышковым организатором. Ядрышко образующие хромосомы кариотипа человека: группы D (№№ 13, 14, 15) и группы G (№№ 21, 22). В этой области расположены гены рРНК. Вторичные перетяжки q-плеча (qh) локализованы в 1, 9, 16 и Y-хромосоме. Функции: Принимают участие в формировании ядрышка, являются местом локализации гетерохроматина. Теломера (t)- концевые участки хромосом, представлены гетерохроматином. Функции: благодаря теломерам хромосомы не слипаются друг с другом. Спутник (S)- небольшой участок хромосомы р-плеча, отделяемый вторичной перетяжкой. Имеется в ядрышко образующих хромосомах. Функция: та же, что и любого участка хромосом. 18. Кариотип и идиограмма человека. Основные показания для исследования кариотипа у человека. Кариотип – совокупность данных о числе, размерах и структуре метафазных хромосом. Каждый вид хромосом в кариотипе, содержащий определенный комплекс генов, представлен двумя гомологами, унаследованными от родителей с их половыми клетками. Двойной набор генов, заключенный в кариотипе,— генотип — это уникальное сочетание парных аллелей генома. В генотипе содержится программа развития конкретной особи. Характеристики: диплоидное (2n) число хромосом, размеры и форма хромосом, наличие перетяжек и спутников, соотношение длин плеч, чередование эу- и гетерохроматина. Идиограмма – графическое изображение хромосом с учетом их абсолютной и относительной длины, центромерного индекса, наличия вторичной перетяжки и спутника. 19. Характеристика Х и У хромосом человека X-хромосома — одна из самых больших во всём наборе 23-х хромосом человека, Y — самая маленькая. Х-хромосома – средняя субметацентрическая , У-хромосома – мелкая акроцентрическая, имеет вторичнуб перетяжку qh. Феномен инактивации хромосомы Х в клетках женского организма на самом деле является тонким фактором регуляции соотношения доз определенных генов, требуемого для воспроизведения нормального фенотипа Характкристика Х- и У-хроматина. Происхождение полового хроматина, и методы его определения, значение в диагностике наследственных заболеваний Х- хроматин (тельце Барра) – это инактивированная и конденсированная одна из двух Х- хромосом соматических интерфазных клеток женщины. В клетках женского эмбриона она инактивируется примерно на 16-20 день развития. Y- хроматин представляет собой блок гетерохроматина в длинном плече Y - хромосомы и выявляется в интерфазных ядрах соматических леток мужчин. В результате в генотипе особей гетерогаметного пола XY гены, расположенные в негомологичных участках Х- и Y-хромосом, встречаются в одной дозе. Препараты полового хроматина можно приготовить, используя клетки слизистой ротовой полости, лейкоцитов, фибробластов кожи и клетки волосяной луковицы, окрашивая их ацетоорсеином или флуоресцентными красителями. У женщин Х-хроматин выявляется с внутренней стороны ядра в виде плотной хорошо окрашенной глыбки, имеющей форму треугольника или овала. Клетки мужчины, как правило, без этой глыбки. женщин половой хроматин (тельце Барра) имеет вид темного тельца, располагающегося у оболочки ядра. 20. Воспроизведение на клеточном уровне. Понятие о жизненном цикле клеток Жизненный (клеточный) цикл клетки (ЖЦК)- период жизни клетки от момента ее рождения (в результате деления материнской клетки) до собственного деления или естественной гибели. Клеточный цикл может включать периоды: Период выполнения клеткой специфических функций. Митотический цикл – процесс подготовки клетки к делению и само деление. Период покоя – ближайшая судьба клетки не определена (стволовые клетки), она может либо начать подготовку к митозу, либо приступить к специализации в определенном функциональном направлении. Основные периоды ЖЦК, утративших способность к делению Гетерокаталетическая интерфаза (транскрибируются гены, контролирующие синтез белков, необходимых для осуществления функции данной клетки) -> смерть Основные периоды ЖЦК клеток, способных к делению Гетерокаталитическая интерфаза(ГКИ) – период жизни клетки, когда она выполняет свои функции -> Митотический цикл (М фаза) -> Автокаталитическая интерфаза (А.К.И.) – период подготовки клетки к делении ->Митоз (<1 часа). В интерфазный период (А.К.И. и Г.К.И.): интерфазные хромосомы при световой микроскопии не выявляются; видны лишь глыбки хроматина (гетерохроматиновые участки хромосом), в том числе Х- и У-хроматин. АВТОКАТАЛИТИЧЕСКАЯ ИНТЕРФАЗА (G1-период) – начальный период интерфазы: транскрибируются гены, контролирующие в основном синтез белков, необходимых для митоза, происходит накопление РНК и белков, в том числе и белков, необходимых для синтеза ДНК, увеличивается количество митохондрий, период длится 12-24 часа. АВТОКАТАЛИТИЧЕСКАЯ ИНТЕРФАЗА (S-период): происходит репликация ДНК, хромосома становится двунитчатой, деконденсированной ДНП (в каждой хромосоме 2 хроматиды) – 2n4c, продолжается также синтез РНК и белков. Длительность S-периода составляет около 5 часов. АВТОКАТАЛИТИЧЕСКАЯ ИНТЕРФАЗА (G2-период): В хромосомы – двунитчатые, деконденсированные ДНП (2n1c), функция ДНП – транскрипция, характеризуется остановкой синтеза ДНК и накоплением энергии, продолжается синтез РНК и белков, формирующий нити веретена деления. Длительность G2-периода составляет 3—6 часов. Митотический цикл - совокупность процессов, происходящих в клетке от одного деления до другого. 21.Цитологическая характеристика периодов и фаз митотичекого цикла Толчком к вступлению клетки в митотический цикл служит изменение соотношения объема ядра и цитоплазмы растущей клетки. Профаза -происходит конденсация и спирализация хромосом, ядерная мембрана растворяется под действием ферментов, ядрышко исчезает; центросома делится на две центриоли, расходятся к полюсам клетки; фосфорилирование отдельных клеточных белков; между полюсами начинает формироваться ахроматиновая фигура (состоит из белка и РНК), которая к концу фазы вытягивается вдоль клетки, становясь веретеном; веретено представляет собой двухполюсную структуру, построенную из микротрубочек и различных белков; хроматиды (сестринские хроматиды) удерживаются вместе центромерои; длительность профазы составляет примерно 30-60 минут. Метафаза - хромосомы располагаются на экваторе веретена, имеют вид толстых образований, плотно свернутых спиралью, прикреплены к нитям веретена центромерои, с посмощью белковых комплексов (кинетофор), связанными с отдельными микротрубочками хроматиды, пока удерживаются вместе, но плечи их уже разъединены. Длительность метафазы составляет 2—10 минут. Анафаза- Хроматиды расходятся к полюсам клетки, конденсированные однонитчатые ДНП (4n4c). Каждая хроматида – самостоятельная хромосома. У каждого полюса по2п2с. В целом в клетке 4п4с. Анафаза длится 2-3 минуты. Репликация хромосомных концов (теломер) требует теломеразы. Телофаза - В телофазе дочерние хромосомы достигают полюсов, вытягиваются и деспирализуются. Хромосомы – однонитчатые ДНП, активность их восстанавливается, при световой микроскопии не выявляются. В ядре каждой дочерней клетки 2п2с. Кинетохорные трубочки исчезают. Образуется ядерная оболочка, вновь появляется ядрышко. Длительность составляет 20-30 минут. Биологическое значение митоза – основной способ деления соматических клеток, обеспечивающий материальную непрерывность генетического материала в популяции клеток, обеспечивает рост организма, регенерацию соматических клеток, а также фазу размножения гаметогоний, из которых впоследствии за счет мейоза формируются половые клетки. |