1. Введение Жизнь на Земле представляет собой форму существования материи
 Скачать 28.03 Mb. Скачать 28.03 Mb.
|

4.3. СтебельСтебель представляет собой ось побега, состоящую из узлов и междоузлий. Основные функции стебля – опорная (несущая) и проводящая. Стебель осуществляет связь между корнями и листьями. В многолетних стеблях обычно откладываются запасные питательные вещества. Молодые стебли, имеющие под эпидермой хлоренхиму, активно участвуют в фотосинтезе. Стебель обычно имеет цилиндрическую форму и характеризуется радиальной симметрией в расположении тканей. Однако в поперечном сечении он может быть не только округлым, но и угловатым – трех-, четырех- или многогранным, ребристым, бороздчатым, иногда совершенно плоским, сплюснутым, или несущим выступающие плоские ребра - крылатым ( рис. 4.27). 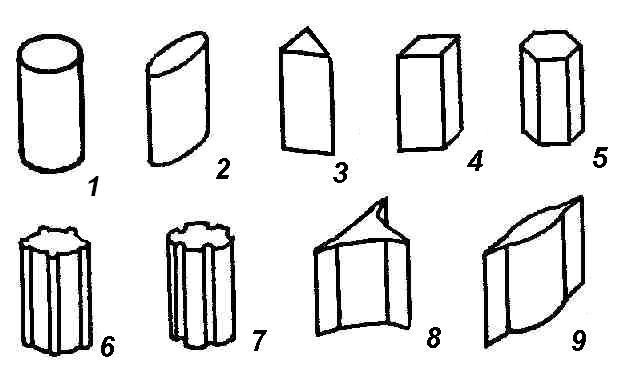 Рис. 4.27. Типы стеблей по форме поперечного сечения : 1 – округлый; 2 – сплюснутый; 3 – трехгранный; 4 – четырехгранный; 5 – многогранный; 6 – ребристый; 7 – бороздчатый; 8, 9 – крылатые. Стебли древесных и травянистых растений резко различаются по продолжительности жизни. Надземные побеги трав сезонного климата живут, как правило, один год; продолжительность жизни побегов определяется продолжительностью жизни стебля. У древесных растений стебель существует много лет. Анатомическое строение стебля соответствует его главным функциям. В стебле развита сложная система проводящих тканей, которая связывает в единое целое все органы растения; наличие механических тканей обеспечивает выполнение опорной функции. Стебель, как и в целом побег, представляет собой «открытую» систему роста, он длительное время нарастает и на нем возникают новые органы. Ткани стебля образуются в результате деятельности сложной системы меристем: верхушечных, боковых и вставочных (рис. 4.28). Первичная структура складывается в результате работы первичных меристем. Инициальные клетки апикальной меристемы сосредоточены в конусе нарастания побега. На апексе побега с правильной периодичностью возникают листовые зачатки, что приводит к раннему вычленению узлов, а развитие междоузлий запаздывает. Часто рост междоузлий и развитие в них постоянных тканей продолжаются длительное время за счет работы остаточных интеркалярных меристем, которые сохраняются у оснований молодых междоузлий. Хорошим примером такого интеркалярного (вставочного) роста может служить стебель злаков, у которого апикальная меристема очень рано расходуется на образование соцветия, и быстрое вытягивание побега обязано именно вставочному росту. 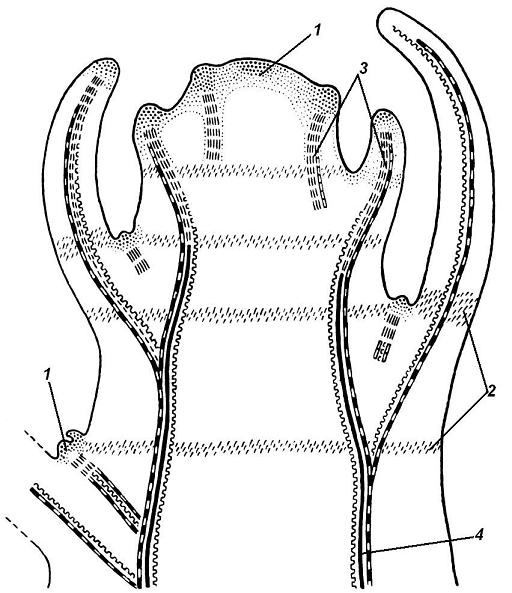 Рис. 4.28. Схема распределения меристем в стебле : 1 – апикальная меристема; 2 – интеркалярная меристема; 3 – прокамбий; 4 – камбий. Самый наружный слой клеток апекса становится протодермой, из которой развивается эпидерма – покровная ткань будущего листа и стебля. На уровне первых листовых бугорков в апикальной меристеме обозначаются тяжи более узких и длинных клеток – это прокамбий, дающий начало первичным проводящим тканям. Прокамбий может возникнуть в виде отдельных пучков или сплошного кольца. По мере дальнейшего роста прокамбий распространяется как в растущий листовой зачаток, так и в стебель, образуя основу будущей проводящей системы побега, связывающей листья и стебли. Остальная часть апекса занята основной меристемой, из которой в дальнейшем формируются паренхимные запасающие и ассимилирующие ткани, а также первичные механические ткани. Основная меристема, находящаяся между протодермой и прокамбием, превращается в первичную кору стебля, а из основной меристемы, располагающейся в центре, образуется сердцевина. Первичная структура стебля у споровых и однодольных растений сохраняется в течение всей жизни. У голосеменных и двудольных внутри прокамбия возникает камбий, который откладывает вторичные проводящие ткани, что приводит к вторичному утолщению стебля. Первичное строение стебля. В стебле, имеющем первичное строение, как и в корне, различают покровную ткань, первичную кору и стелу (осевой, или центральный цилиндр) ( рис. 4.29). Покровной тканью является эпидерма типичного строения. В состав первичной коры входит основная паренхима, а также механические, выделительные и некоторые другие ткани. Из механических тканей чаще встречается колленхима, она образует либо сплошной цилиндр, либо имеет вид отдельных тяжей, располагающихся обычно вдоль выступов – ребер стебля (рис. 4.29 ). Сразу под колленхимой или эпидермой, если колленхима отсутствует, в условиях, благоприятных для фотосинтеза, располагается хлоренхима. Она может образовывать с колленхимой или склеренхимой чередующиеся полосы, проходящие вдоль стебля. Граница между корой и стелой выражена гораздо менее четко, нежели в корне, так как внутренний слой коры – эндодерма – не имеет характерных признаков, присущих эндодерме корня. Нередко в нем откладываются крахмальные зерна, играющие, по-видимому, роль статолитов, подобно крахмальным зернам корневого чехлика. Поэтому эндодерму в стебле называют крахмалоносным влагалищем. Эндодерма типичного для корня строения иногда развивается в корневищах однодольных растений (ландыш) ( рис. 4.48). 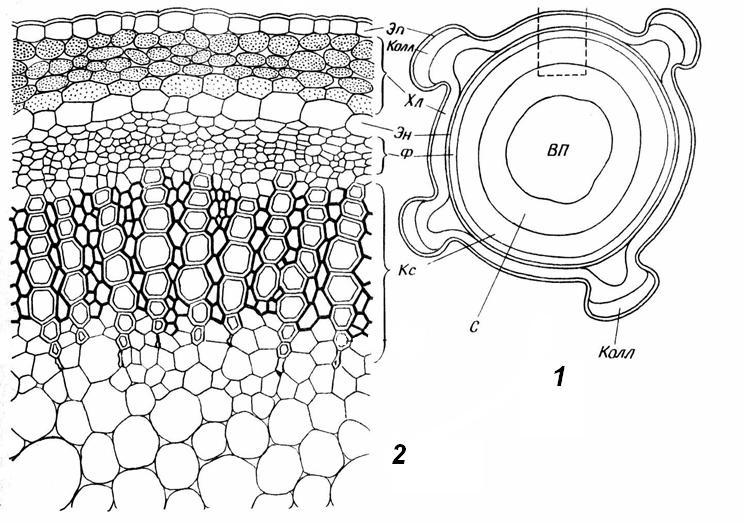 Рис. 4.29. Поперечный срез стебля подмаренника мягкого (первичное строение): 1 – общая схема; 2 – часть среза при большем увеличении; ВП – воздушная полость; Колл – колленхима; Кс – ксилема; С – сердцевина; Ф - флоэма; Хл – хлоренхима; Эн – эндодерма (крахмалоносное влагалище); Эп – эпидерма. Наиболее сложное строение имеет стела. У голосеменных и двудольных растений первичные проводящие ткани чаще всего разделены на проводящие пучки, которые на поперечном срезе располагаются в виде кольца; между пучками находятся клетки паренхимы, которые составляют первичные сердцевинные лучи. Иногда разделение на пучки неотчетливо или совсем незаметно (рис. 4.29) . Первичная ксилема лежит рядом с сердцевиной, а первичная флоэма – кнаружи от ксилемы, рядом с корой. Флоэма и ксилема в стебле развиваются навстречу друг другу. Кроме наружной флоэмы, иногда возникает и внутренняя, которая находится между ксилемой и сердцевиной. Внутренняя флоэма может входить в состав пучков (тыква), располагается в виде самостоятельных тяжей (томат) или же образует сплошной слой (олеандр). Наружная часть стелы носит название перицикла. Перицикл может быть представлен клетками основной паренхимы и склеренхимными волокнами. Волокна образуют сплошной слой или собраны около проводящих пучков; в последнем случае говорят о сосудисто-волокнистых пучках. Природа склеренхимы, возникающей между корой стебля и проводящими элементами флоэмы, остается не вполне ясной. Доказано, что она не всегда имеет перициклическое происхождение, во многих случаях волокна формируются в первичной флоэме ( рис. 4.36). Внутрь от проводящих тканей располагается сердцевина, состоящая из неспециализированной паренхимы. Иногда в ней откладываются запасные вещества, или могут быть рассеяны идиобласты с таннидами, слизями и др. Часто в сердцевине образуется воздушная полость. Периферическая часть сердцевины, примыкающая к ксилеме, состоит из более мелких и плотно расположенных клеток и называется перимедуллярной зоной. Характерной особенностью стебля является расположение механических тканей на периферии органа и их отсутствие в центре. Это связано с тем, что стебель, в отличие от корня, развивается в воздушной среде и испытывает в основном динамические нагрузки (порывы ветра, удары дождевых капель, вытаптывание животными и т.д.). Поэтому стебель имеет конструкцию полой упругой пружины, способной возвращаться в исходное состояние после снятия нагрузки. Корню, окруженному почвой, не грозит опасность изгиба и излома. Корень «заякоривает» растение в почве и противодействует напряжениям, стремящимся выдернуть его. Соответственно этому механические элементы размещены в самом центре корня. У однодольных растений отсутствует камбий, и первичное строение сохраняется в течение всей жизни. Характерной особенностью стеблей однодольных является всегда пучковое строение, причем закрытые проводящие пучки располагаются без видимого порядка по всему поперечному сечению (рис. 4.30 ). 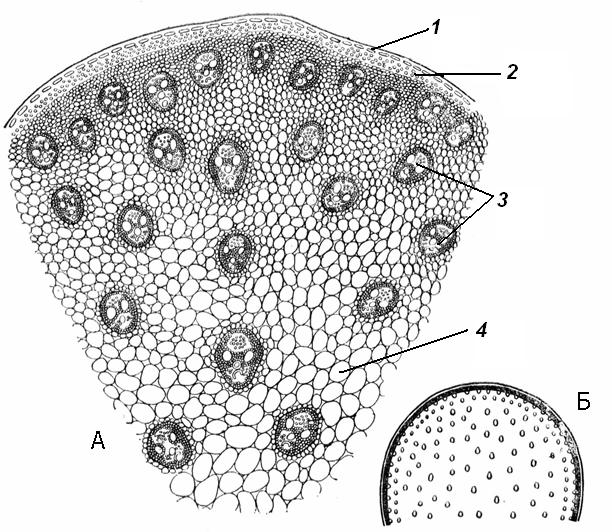 Рис. 4.30. Строение стебля однодольного растения (кукуруза): А – поперечный срез; Б – общая схема; 1 – эпидерма; 2 – склеренхима; 3 – закрытые коллатеральные пучки; 4 – основная паренхима. К основным типам строения стебля однодольных относят стебель, не имеющий полости, (рис. 4.30) и стебель с одной большой центральной полостью (соломина злаков) (рис. 4.31). В последнем случае пучки располагаются по периферии органа. Высокая прочность стебля достигается обильным развитием склеренхимы. Первичная кора у однодольных часто не выражена. 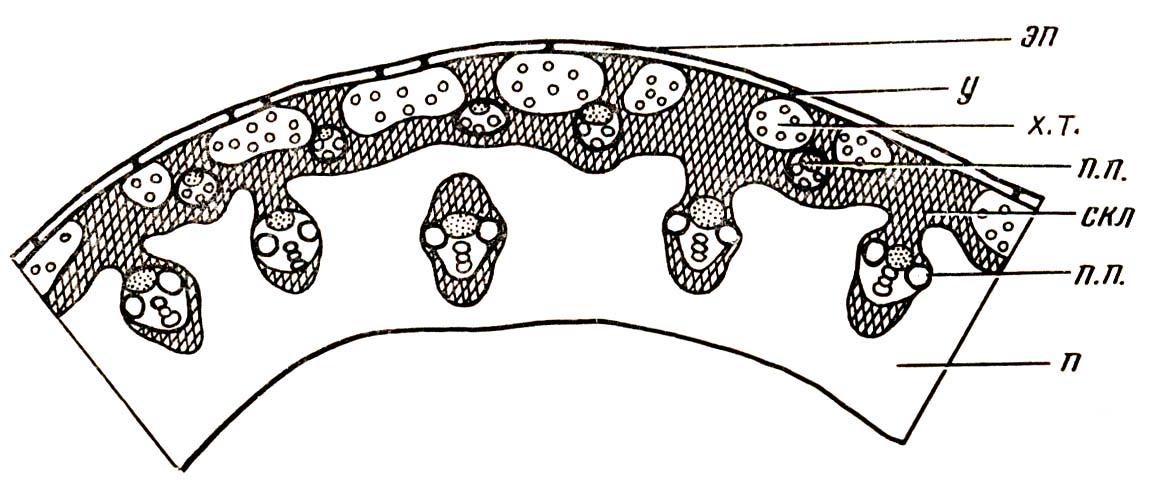 Рис. 4.31. Схема строения соломины ржи : п – паренхима; п. п. – закрытые коллатеральные проводящие пучки; скл – склеренхима; у – устьице; х. т. – хлорофиллоносная ткань; эп – эпидерма. Наряду с коллатеральными проводящими пучками (рис. 4.32), в стеблях однодольных растений довольно обычны концентрические амфивазальные (центрофлоэмные) пучки ( рис. 4.33). 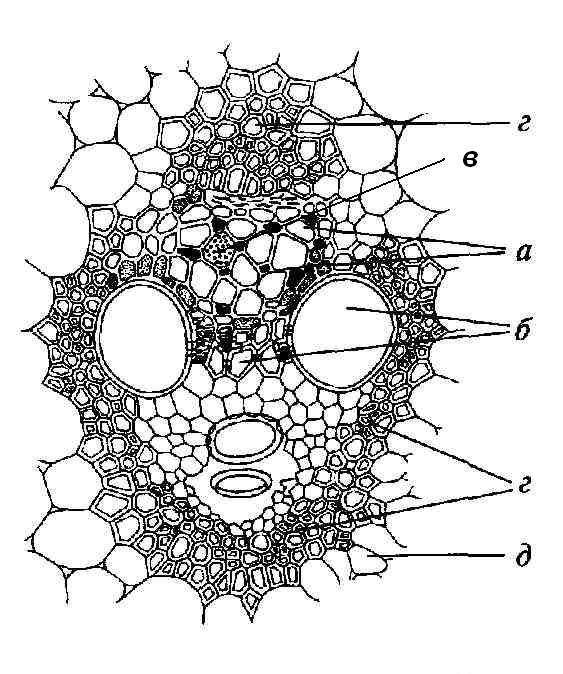 Рис. 4.32. Закрытый коллатеральный пучок стебля кукурузы : а – первичная флоэма; б – первичная ксилема; в – ситовидная пластинка; г – склеренхимная обкладка; д – основная паренхима стебля, окружающая пучок. Стебли однодольных могут достигать значительной толщины за счет первичного утолщения - деления и растяжения клеток апикальной меристемы и тех первичных тканей, которые из нее возникают (пальмы). У некоторых древесных однодольных (драцены, юкки, алоэ) отмечено вторичное утолщение стеблей, которое происходит иначе, чем у двудольных (рис. 4.33 ). 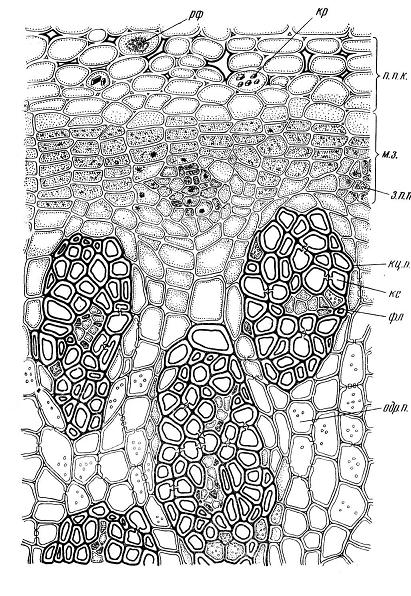 Рис. 4.33. Строение стебля драцены в зоне вторичного утолщения: з. п. п. – закладывающийся проводящий пучок; кр – кристаллы оксалата кальция; кс – ксилема; кц. п. – концентрический пучок; м. з. – меристематическая зона; одр. п. – одревесневшая паренхима; п. п. к. – паренхима первичной коры; рф – рафиды в поперечном разрезе; фл – флоэма. На периферии стебля сохраняется зона клеток меристемы (зона вторичного утолщения). Большинство клеток, откладываемых меристемой внутрь, превращается в клетки основной паренхимы, часть дифференцируется в закрытые проводящие пучки. Клетки, откладываемые наружу, образуют покровную ткань, сходную с перидермой. Рассмотренное вторичное утолщение служит подтверждением закона необратимости эволюционного процесса: однодольные произошли от предков, обладавших нормальным камбиальным утолщением, но, потеряв камбий, уже не смогли восстановить его вновь. Вторичное строение стебля. У большинства двудольных и голосеменных растений довольно рано возникают вторичные изменения анатомической структуры стебля. Они связаны, главным образом, с активностью боковой вторичной меристемы – камбия и, отчасти, - другой вторичной меристемы – феллогена. Камбий возникает из остатков прокамбия, на границе первичных ксилемы и флоэмы. Он состоит из тонкостенных клеток, заостренных на концах и вытянутых вдоль оси стебля (рис. 4.34 ). Плоские широкие стороны каждой клетки обращены внутрь, в сторону ксилемы, и наружу, в сторону флоэмы. Деление клетки происходит тангенциально, т. е. параллельно плоским сторонам. После каждого деления одна из дочерних клеток остается в составе инициалей, другая может еще разделиться два-три раза, но судьба ее производных предрешена. Если они расположены внутрь от инициальной клетки, то превращаются в элементы ксилемы, если наружу, то в элементы флоэмы. Обычно внутрь откладывается большее число производных, чем наружу, и ксилема нарастает быстрее флоэмы. Поскольку дифференциация производных камбия происходит не сразу, формируется камбиальная зона, в составе которой имеется только один слой инициалей, способных сохранять свою активность очень долго. 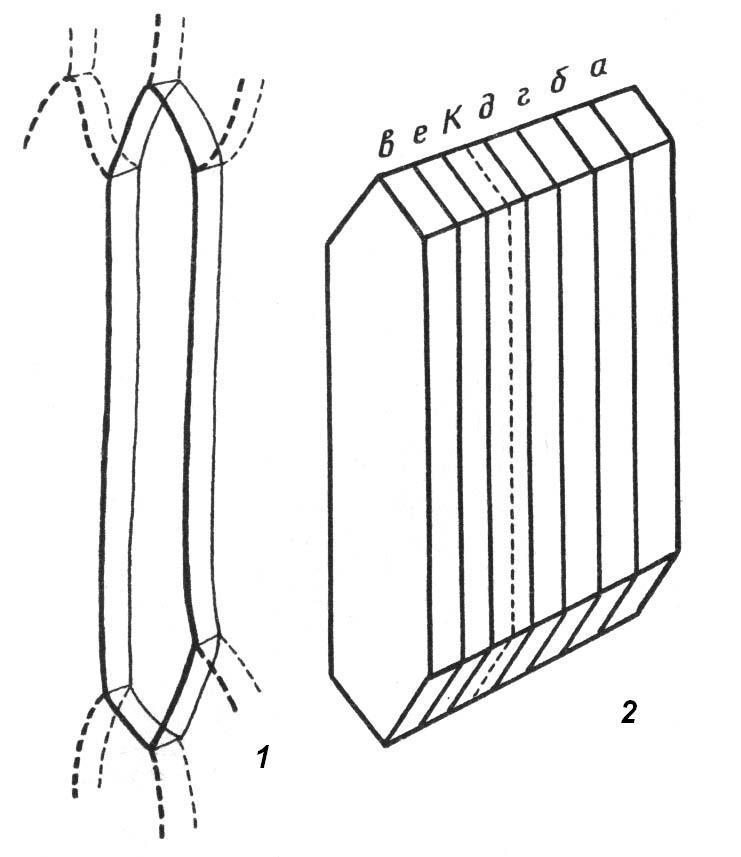 Рис. 4.34. Схема строения и работы камбия : 1 – форма инициальной веретеновидной клетки камбия; 2 – порядок двустороннего отложения элементов (а….е); К – инициальная клетка камбия (пунктиром показана плоскость следующего деления). Вторичную ксилему называют древесиной, а вторичную флоэму – лубом. По мере утолщения стебля камбиальный слой растягивается, поэтому время от времени инициальные клетки делятся радиально, увеличивая общую поверхность камбиального слоя. Кроме сильно вытянутых (веретеновидных) клеток, камбий содержит группы коротких (лучевых) инициальных клеток, дающих начало лубодревесинным, или вторичным сердцевинным лучам. В районах с выраженным сезонным климатом в деятельности камбия наблюдаются периоды покоя, совпадающие с зимним понижением температуры или с засушливым периодом. Возникновение камбия и интенсивность его работы неодинаковы у разных растений. Камбий может возникать с самого начала как непрерывное кольцо в сплошном слое прокамбия и затем длительно откладывать сплошные слои вторичных проводящих тканей (рис. 4.35 ). В этом случае формируется непучковое строение стебля. Проводящие ткани имеют вид полых цилиндров, вставленных один в другой. 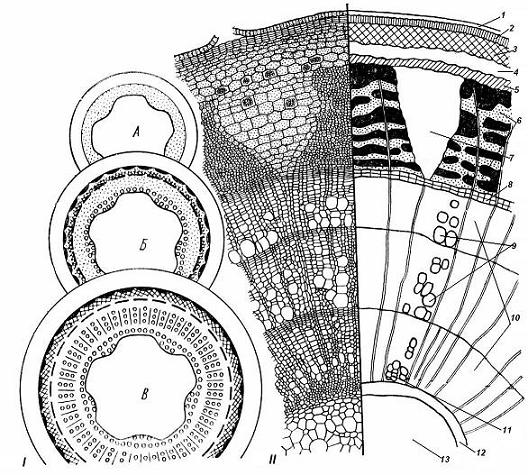 Рис. 4.35. Стебель липы на поперечном срезе (II) и схема строения на разных уровнях (I): А – срез на уровне появления прокамбия; Б – срез на уровне появления камбия; В – срез на уровне сформированной структуры; 1 – остатки эпидермы; 2 – перидерма; 3 – колленхима; 4 – паренхима; 5 – эндодерма (3 – 5 – первичная кора); 6 – вторичная флоэма; 7 – первичный сердцевинный луч; 8 – камбий; 9 – годичные кольца; 10 – вторичная ксилема; 11 – первичная ксилема; 12 – перимедуллярная зона; 13 – сердцевина. У других растений в прокамбиальных пучках сначала возникает пучковый камбий. Затем между разобщенными прослойками пучкового камбия возникают перемычки межпучкового камбия, после чего образовавшийся непрерывный камбиальный слой откладывает сплошные слои вторичных тканей. При этом также формируется непучковое строение органа. Непучковый тип строения, прежде всего, характерен для многолетних, длительно утолщающихся стеблей деревьев и кустарников, но встречается и у травянистых растений ( рис. 4.36). 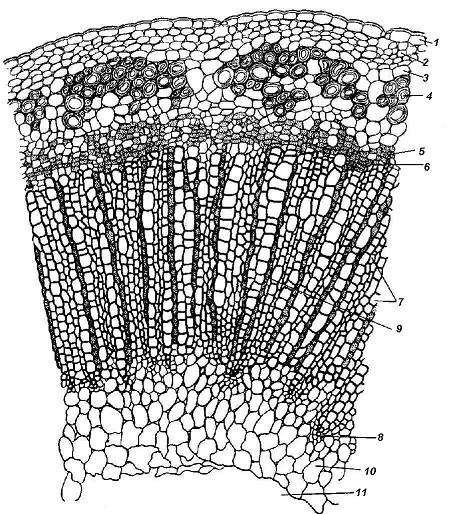 Рис. 4.36. Поперечный срез стебля льна (непучковое строение): 1 – эпидерма; 2 – паренхима первичной коры; 3 – эндодерма; 4 – первичные лубяные волокна; 5 – флоэма; 6 – камбий; 7 – вторичная ксилема; 8 – первичная ксилема; 9 – сердцевинный луч; 10 – паренхима сердцевины; 11 – воздушная полость. В стеблях травянистых двудольных растений камбий функционирует непродолжительное время. Как правило, вторичные проводящие ткани образуются только за счет пучкового камбия, и стебель сохраняет в течение всей жизни пучковое строение (рис. 4.37). Межпучковый камбий образует только механические элементы, составляющие в совокупности с пучками трубчатую скелетную конструкцию, или тонкостенную паренхиму, неотличимую от паренхимы коры и сердцевины (рис. 4.37 ). 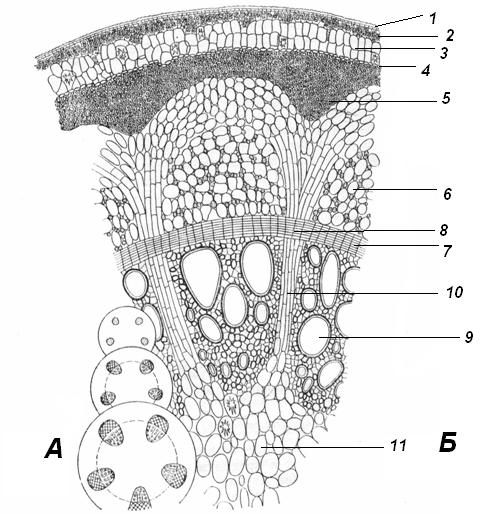 Рис. 4.37. Стебель кирказона на поперечном срезе (Б) и схема строения на разных уровнях (А): 1 – эпидерма; 2 – колленхима; 3 – паренхима первичной коры; 4 – эндодерма; 5 – склеренхима перицикла; 6 – флоэма; 7 – пучковый камбий; 8 – межпучковый камбий; 9 – ксилема; 10 – сердцевинный луч; 11 – сердцевина. Стебли некоторых травянистых двудольных имеют так называемое переходное строение от пучкового к непучковому типу (рис. 4.38 ). В этом случае первоначально формируются проводящие пучки, и функционирует только пучковый камбий. Межпучковый камбий возникает позднее и начинает откладывать элементы новых проводящих пучков: ксилему внутрь от камбия и флоэму кнаружи. Постепенно новые и старые пучки разрастаются и сливаются в сплошной слой проводящих тканей. 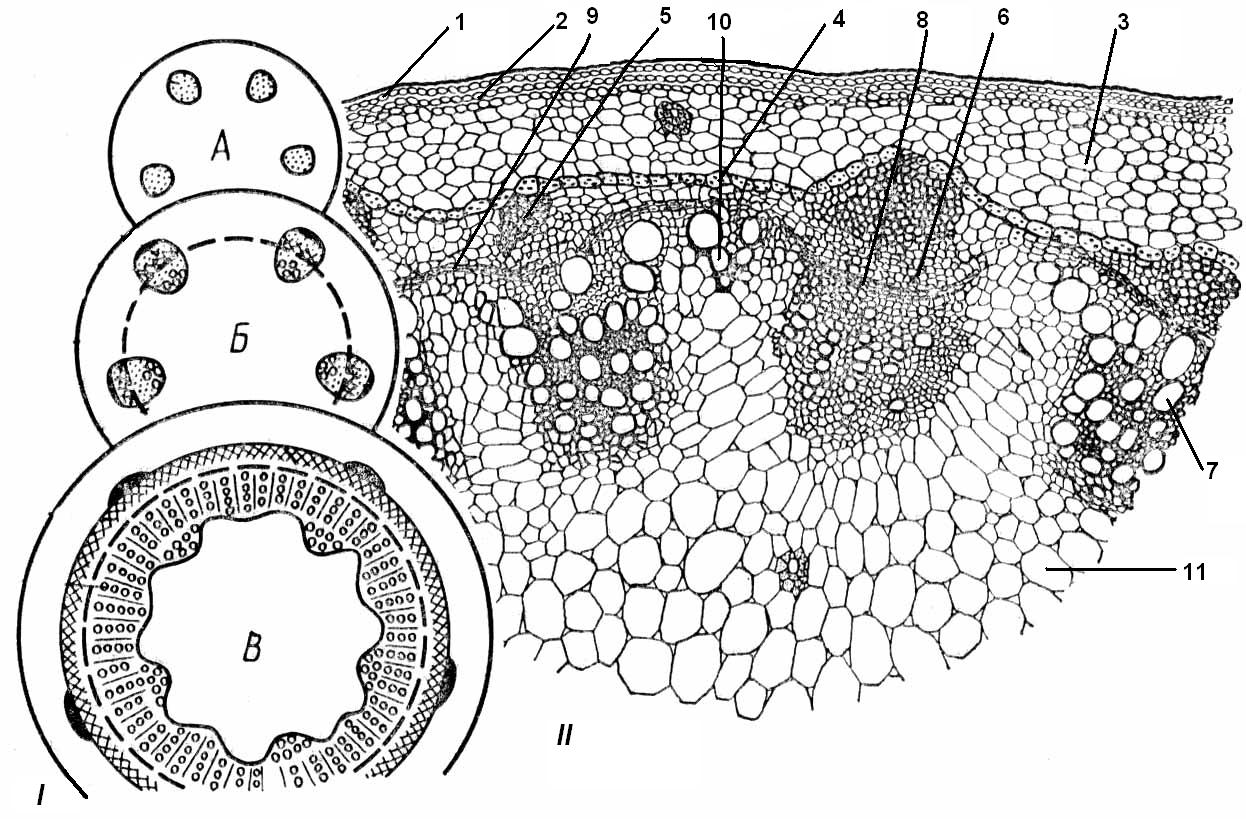 Рис. 4.38. Стебель подсолнечника на поперечном срезе ( II) и схема строения на разных уровнях (I): А – срез на уровне появления прокамбия; Б – срез на уровне появления камбия; В – срез на уровне сформированной структуры: 1 – эпидерма; 2 – колленхима; 3 – паренхима первичной коры; 4 – эндодерма (крахмалоносное влагалище); 5 – склеренхима; 6 – флоэма; 7 – ксилема; 8 – пучковый камбий; 9 – межпучковый камбий; 10 – пучок из межпучкового камбия; 11 – паренхима сердцевины. При пучковом типе строения у двудольных растений проводящие пучки всегда открытые, чаще коллатеральные (рис. 4.37, рис. 4.38), у некоторых растений (тыква, огурец) – биколлатеральные (рис. 4.39) ; располагаются пучки на поперечном срезе, в отличие от однодольных, по кругу (кольцом). 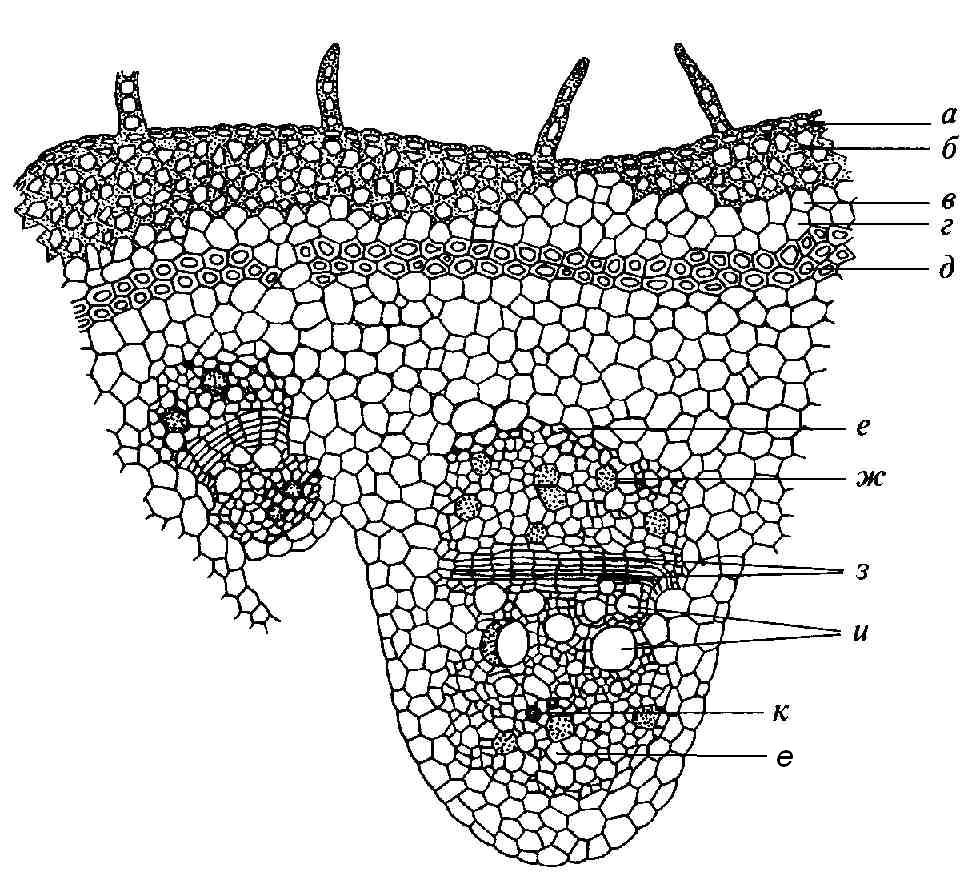 Рис. 4.39. Фрагмент поперечного среза стебля тыквы : а – эпидерма; б – колленхима; в – паренхима первичной коры; г – эндодерма; д – склеренхима перицикла; е – первичная флоэма; ж – вторичная флоэма; з – камбиальная зона; и – вторичная ксилема; к - первичная ксилема. Очень редко у двудольных растений с тонким слабым стеблем камбий вообще не образуется, и проводящие пучки, состоящие из первичных проводящих тканей, включены в однородную первичную паренхиму ( рис. 4.40). 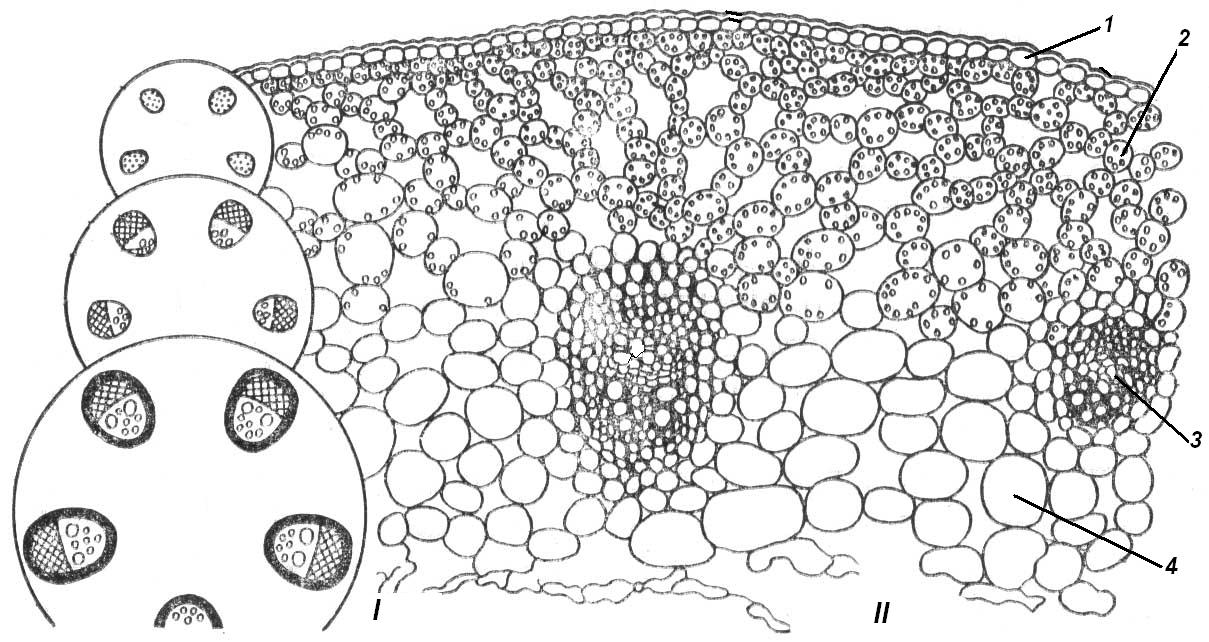 Рис. 4.40. Стебель лютика на поперечном срезе ( II) и схема строения на разных уровнях (I): 1 – эпидерма; 2 – первичная кора (хлоренхима и аэренхима); 3 – коллатеральный закрытый пучок; 4 – паренхима сердцевины. У древесных двудольных и хвойных растений камбиальное утолщение продолжается долгие годы. Секвойя образует ствол толщиной свыше10 м. Центр ствола занят вторичной ксилемой (древесиной), составляющей около 0,9 всего объема органа (рис. 4.41 ). На поверхности древесины находится тончайший слой камбия, а кнаружи от него – вторичная кора. В состав вторичной коры входят: вторичная флоэма (луб), остатки первичной флоэмы и первичной коры, а также перидерма, сменившая эпидерму в качестве покровной ткани. Позднее первичные ткани становятся совершенно неразличимыми, а из наружных слоев вторичной коры формируется третичная покровная ткань – корка. Мертвую корку часто называют наружной корой, а живую часть коры между камбием и самым внутренним слоем феллогена – внутренней корой.  Рис. 4.41. Схема строения ствола дуба (темная зона в центре – ядро, светлая часть древесины - заболонь). Массовый транспорт веществ в стволе идет по молодым слоям луба и древесины, расположенным около камбия. Луб обычно теряет способность к транспорту через год (после перезимовывания), древесина – через несколько лет. На смену стареющим тканям камбий откладывает новые слои, поэтому вблизи камбия всегда сохраняется тонкий слой живых жизнедеятельных тканей. Основная масса ствола состоит из мертвых клеток и не принимает непосредственного участия в транспорте веществ. Тем не менее, отмершие ткани имеют громадное значение: древесина поддерживает колоссальную тяжесть кроны, а мертвые ткани коры защищают внутренние живые ткани. Вторичные проводящие ткани имеют сложное строение и образованы различными типами клеток. В состав древесины двудольных растений входят следующие основные элементы: сосуды, трахеиды, древесинные волокна (волокна либриформа), клетки вертикальной (древесинной) паренхимы (рис. 4.42 ), а также живые клетки древесинных лучей (лучевой, или древесной паренхимы) ( рис. 4.43). 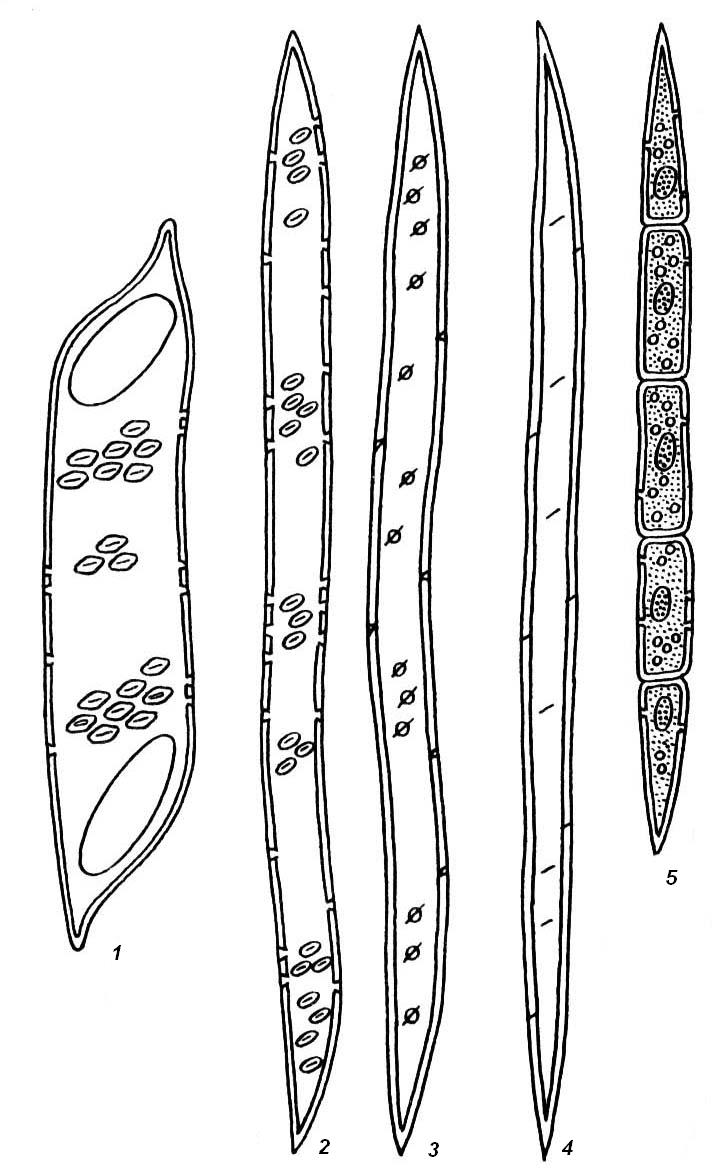 Рис. 4.42. Элементы, входящие в состав вертикальной системы древесины: 1 – членик сосуда; 2, 3 – трахеиды; 4 – древесинное волокно (волокно либриформа); 5 – тяж древесинной паренхимы. Главная масса древесины составлена клетками, лишенными живого содержимого (сосуды, трахеиды, волокна). Среди них распределены живые элементы лучевой и вертикальной паренхимы, образующие связную систему, по которой передвигаются запасные вещества. В этих же живых клетках откладываются запасные вещества, которые весной превращаются в сахара и с водным током поднимаются по водопроводящим элементам древесины вверх, к молодым, энергично растущим побегам. Установлено, что хотя водопроводящими элементами являются не имеющие в зрелом состоянии протопластов сосуды и трахеиды, они могут выполнять свою функцию лишь до тех пор, пока в древесине содержатся живые элементы. 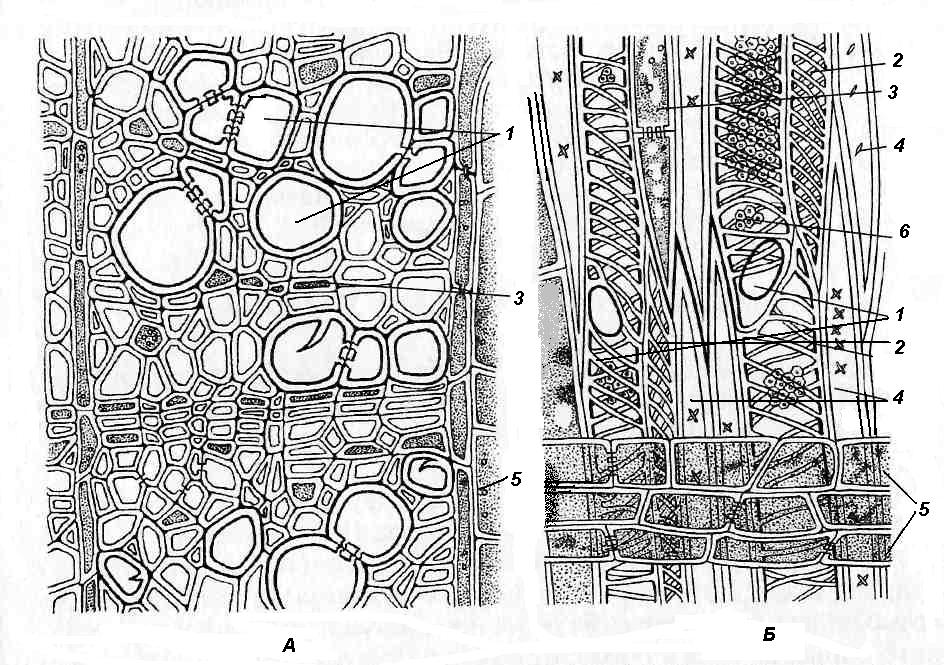 Рис. 4.43. Строение древесины липы на поперечном (А) и радиальном (Б) срезах: 1 – сосуды; 2 – трахеиды; 3 – древесинная паренхима; 4 – древесинные волокна (волокна либриформа); 5 – древесинный луч; 6 – окаймленная пора. Горизонтальные ленточки древесинных лучей образуются лучевыми инициалями камбия. Так как соседние лучи не параллельны, а расходятся по радиусам, то при утолщении ствола наружные участки лучей удаляются друг от друга. Увеличение расстояния между ними ухудшает деятельность тех клеток, которые оказываются слишком удаленными от лучей. Поэтому время от времени между старыми лучами появляются новые. Все элементы расположены в древесине строго упорядоченно. Камбий работает с определенной ритмичностью, зависящей от сезонов года. Наивысшей активностью камбий обладает весной, когда растут молодые побеги и развертываются листья. В это время камбий откладывает преимущественно водопроводящие элементы с широкими полостями и тонкими стенками. Летом активность камбия снижается. Он откладывает преимущественно узкие элементы с толстыми стенками (волокна и толстостенные трахеиды). К осени камбий совсем прекращает работу. После осенне-зимнего периода покоя работа камбия возобновляется следующей весной. Так как переход от весенней (ранней) древесины к летней (поздней) древесине постепенный, а от летней к весенней – резкий, внезапный, то в древесине возникают годичные слои с отчетливыми границами. На поперечном срезе эти слои видны как годичные кольца (рис. 4.35 ). По числу годичных слоев можно установить возраст побега. Однако многие деревья, растущие в бессезонном климате влажных тропиков, не образуют заметных годичных слоев. В районах умеренного климата годичные слои в неблагоприятных условиях могут не возникать или удваиваться, например в тех случаях, когда листья уничтожались вредителями и затем отрастали вновь. На ширину годичных слоев влияют условия произрастания и погодные условия. В благоприятные годы образуются широкие слои, а в неблагоприятные – узкие. Сопоставляя чередование слоев в образцах древесины, можно вычертить график, отражающий колебания погодных условий за многие века. Возрастные изменения, приводящие к отмиранию живых элементов древесины, включают морфологические изменения и образование ряда веществ. У ряда растений образуются тилы. Тила представляет собой вырост живой клетки, внедрившейся через пору в полость сосуда (рис. 4.44 ). Тилы закупоривают сосуды и прекращают передвижение по ним веществ. Часто стенки тил сильно утолщаются, а внутри накапливаются смолы и дубильные вещества, препятствующие деятельности грибов – разрушителей древесины. Тилы могут выполнять также запасающую функцию. 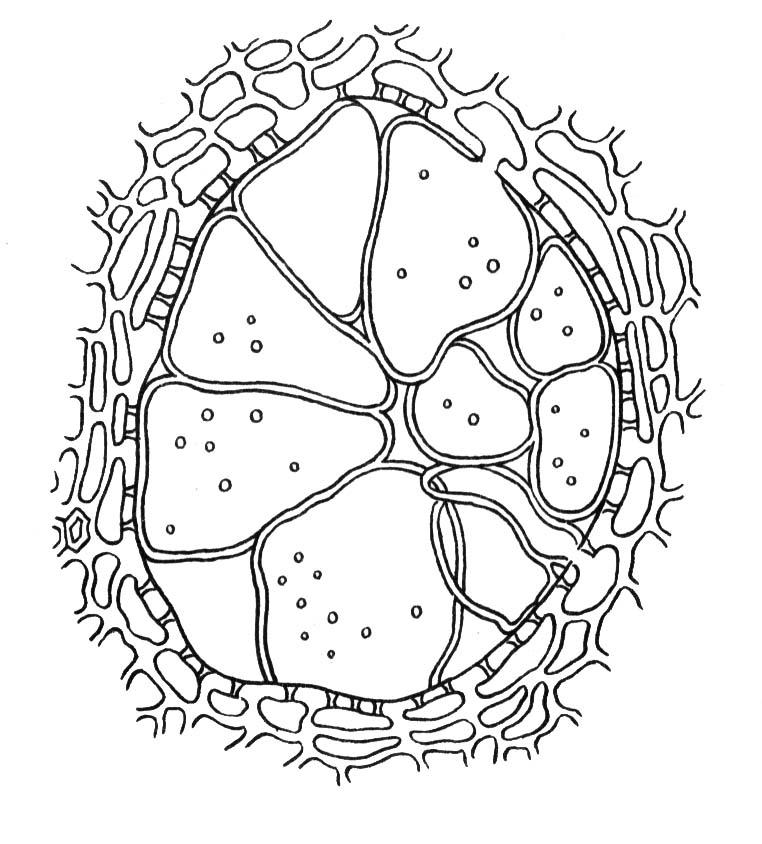 Рис. 4.44. Сосуд, заполненный тилами. Молодую древесину, лежащую около камбия, называют заболонной или просто зaболонью. Внутрь от заболони расположена древесина, которая имеет меньшую влажность и, по-видимому, почти не принимает участия в проведении воды. Если эта древесина по виду не отличается от заболони, ее называют спелой. Соответственно, деревья, имеющие спелую древесину (осина, бук, ель, груша), называют спелодревесными. У ряда других растений (сосна, дуб, ясень, вяз) внутренняя древесина (ядро) имеет более темный по сравнению с заболонью цвет за счет образования дубильных, красящих и смолистых веществ (рис. 4.41). Такие древесины называют ядровыми. Особенности анатомического строения древесины определяют ее технологические и декоративные свойства. Наличие сосудов – важнейший отличительный признак древесины покрытосеменных растений. У споровых и голосеменных водопроводящие элементы представлены только трахеидами. У хвойных растений основную массу древесины составляют трахеиды (рис. 4.45 ). Они располагаются правильными радиальными рядами, каждый ряд отложен одной клеткой камбия. Ранняя древесина представлена широкопросветными тонкостенными трахеидами с хорошо выраженными окаймленными порами на радиальных стенках. Эти трахеиды выполняют преимущественно водопроводящую функцию. В поздней древесине трахеиды имеют толстые стенки и узкие полости, они выполняют преимущественно механическую функцию. Между годичными слоями граница очень резкая. 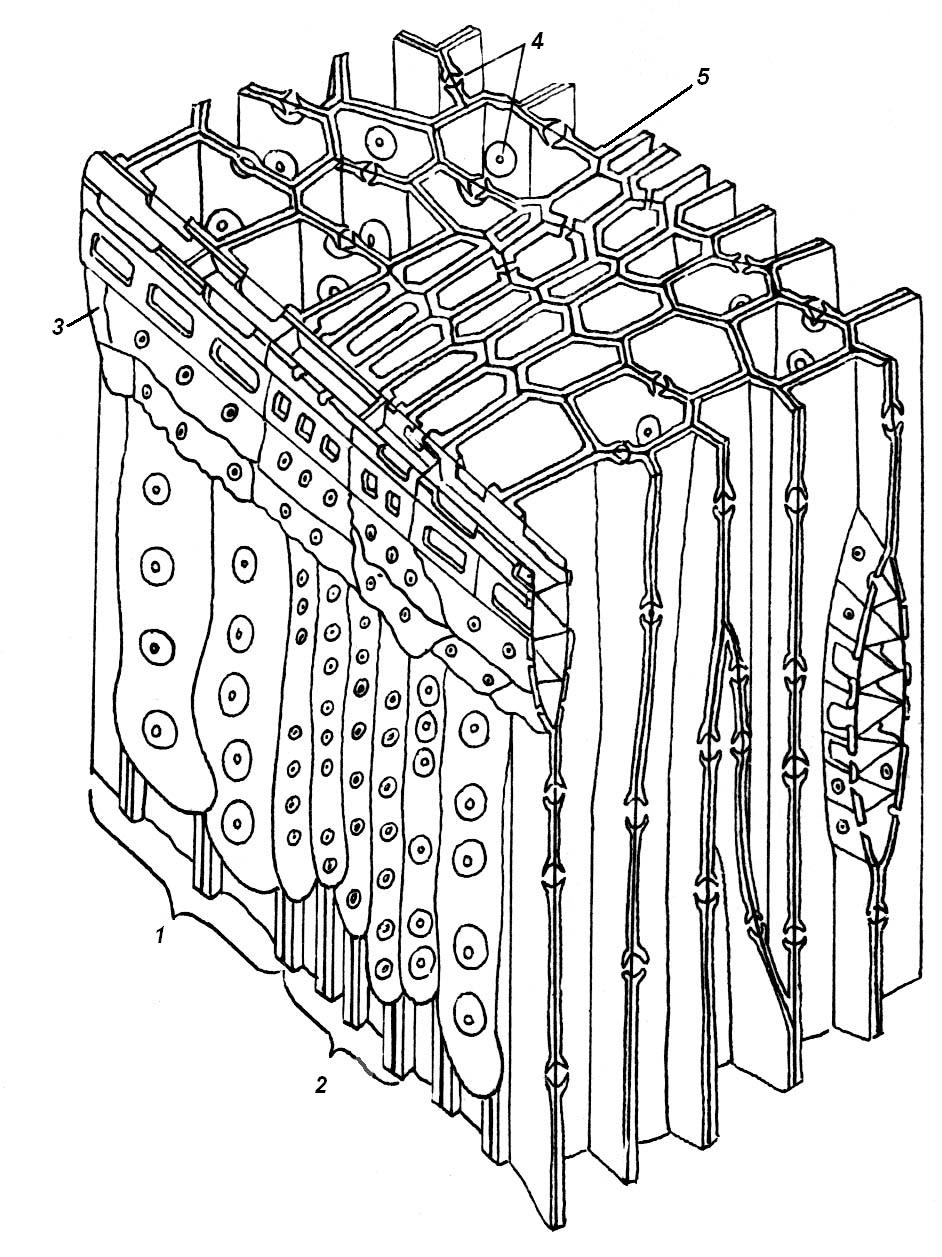 Рис. 4.45. Схема строения древесины хвойного растения : 1 – ранняя древесина; 2 - поздняя древесина; 3 – луч; 4 – окаймленные поры; 5 – граница годичного кольца. Среди трахеид проходят лучи. Каждый луч представляет собой ленточку, проходящую радиально из древесины сквозь камбий в кору. Древесинный луч составлен клетками двух типов. Верхний и нижний ярусы луча состоят из мертвых клеток, вытянутых вдоль луча. На их стенках имеются мелкие окаймленные поры. Эти клетки называются лучевыми трахеидами. Их функция – проведение воды в радиальном направлении. Клетки луча, расположенные в среднем ярусе, остаются живыми, по ним в радиальном направлении передвигаются пластические вещества. У многих хвойных в древесине имеется целая система вертикальных и горизонтальных смоляных каналов схизогенного типа. Полости каналов заполнены смолой, которая при повреждении вытекает наружу, ее летучие фракции испаряются, смола затвердевает и закрывает рану. В целом, древесина покрытосеменных растений имеет значительно более сложную и разнообразную структуру, чем древесина голосеменных. Показано, что по стволам лиственных древесных растений вода поднимается гораздо быстрее, чем по стволам хвойных. Полагают, что структурная эволюция древесины была одной из причин быстрого расцвета покрытосеменных и расселения их на обширных пространствах и в различных условиях обитания. Вторичный луб, подобно древесине, состоит из элементов двух систем – вертикальной и горизонтальной (радиальной). К вертикальной системе принадлежат ситовидные трубки с сопровождающими клетками, вертикальные тяжи лубяной паренхимы и лубяные волокна. Горизонтальные элементы представлены лубяными лучами (рис. 4.46). Лубяные волокна чаще всего залегают в виде прослоек так называемого твердого луба, между которыми находятся живые тонкостенные элементы мягкого луба. В состав мягкого луба входят ситовидные элементы с сопровождающими клетками и лубяная паренхима. 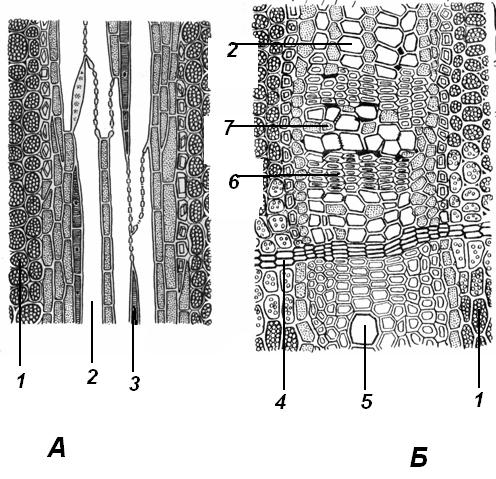 Рис. 4.46. Строение вторичного луба винограда : А – продольный тангенциальный срез; Б – поперечный срез; 1 – сердцевинный луч; 2 – ситовидная трубка; 3 – сопровождающая клетка; 4 – камбий; 5 – древесина; 6 – лубяные волокна (твердый луб); 7 – мягкий луб. У хвойных растений нет ситовидных трубок с сопровождающими клетками, транспорт ассимилятов осуществляют более примитивные проводящие элементы - ситовидные клетки, обладающие в зрелом состоянии ядрами. На судьбу вторичного луба сильное влияние оказывает увеличение толщины ствола. По мере увеличения массы древесины кора отодвигается от центра и при этом испытывает деформацию в двух направлениях – растягивается по окружности и сдавливается в радиальном направлении. Сжатие луба в радиальном направлении приводит к тому, что он быстро теряет способность к проведению ассимилятов. Ситовидные элементы сдавливаются и отмирают. Лишь у некоторых деревьев они способны функционировать несколько лет (липа). Зона коры, проводящая вещества в вертикальном направлении, таким образом, ничтожно мала – около 1 мм. Часть паренхимных клеток луба может превращаться в толстостенные склереиды, что повышает прочность коры. Во избежание разрывов при растягивании в тангенциальном направлении тонкостенные живые клетки первичных сердцевинных лучей подвергаются дилатации (разрастаются). Такие лучи имеют вид треугольников, обращенных вершинами к камбию (рис. 4.35 ). Строение корневищ. Корневище представляет собой видоизмененный подземный побег, и, в целом, анатомическая структура корневища соответствует структуре надземного стебля данного растения. К особенностям строения корневищ можно отнести слабое развитие или полное отсутствие механических тканей и большое количество запасающей паренхимы. Эти особенности объясняются запасающей функцией корневища и тем, что, в отличие от ортотропного стебля, ему не нужно поддерживать в пространстве боковые побеги и листья. Поскольку запасающая паренхима располагается обычно в сердцевине и в первичной коре, последняя в корневищах развита, как правило, лучше, чем в надземных стеблях соответствующих растений. Корневища однодольных растений, также как и стебли, имеют всегда пучковое строение. Пучки разбросаны по всему поперечному сечению, они закрытые, коллатеральные (рис. 4.47) и (или) амфивазальные (центрофлоэмные) (рис. 4.48). Концентрические проводящие пучки в корневищах встречаются чаще, чем в надземных стеблях. 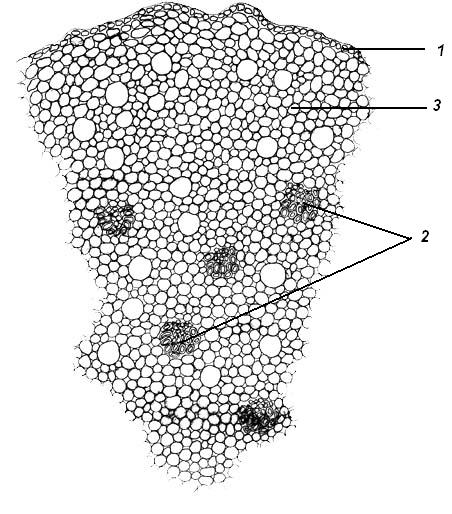 Рис. 4.47. Поперечный срез корневища купены : 1 – эпидерма; 2 – закрытые коллатеральные пучки; 3 – запасающая паренхима. 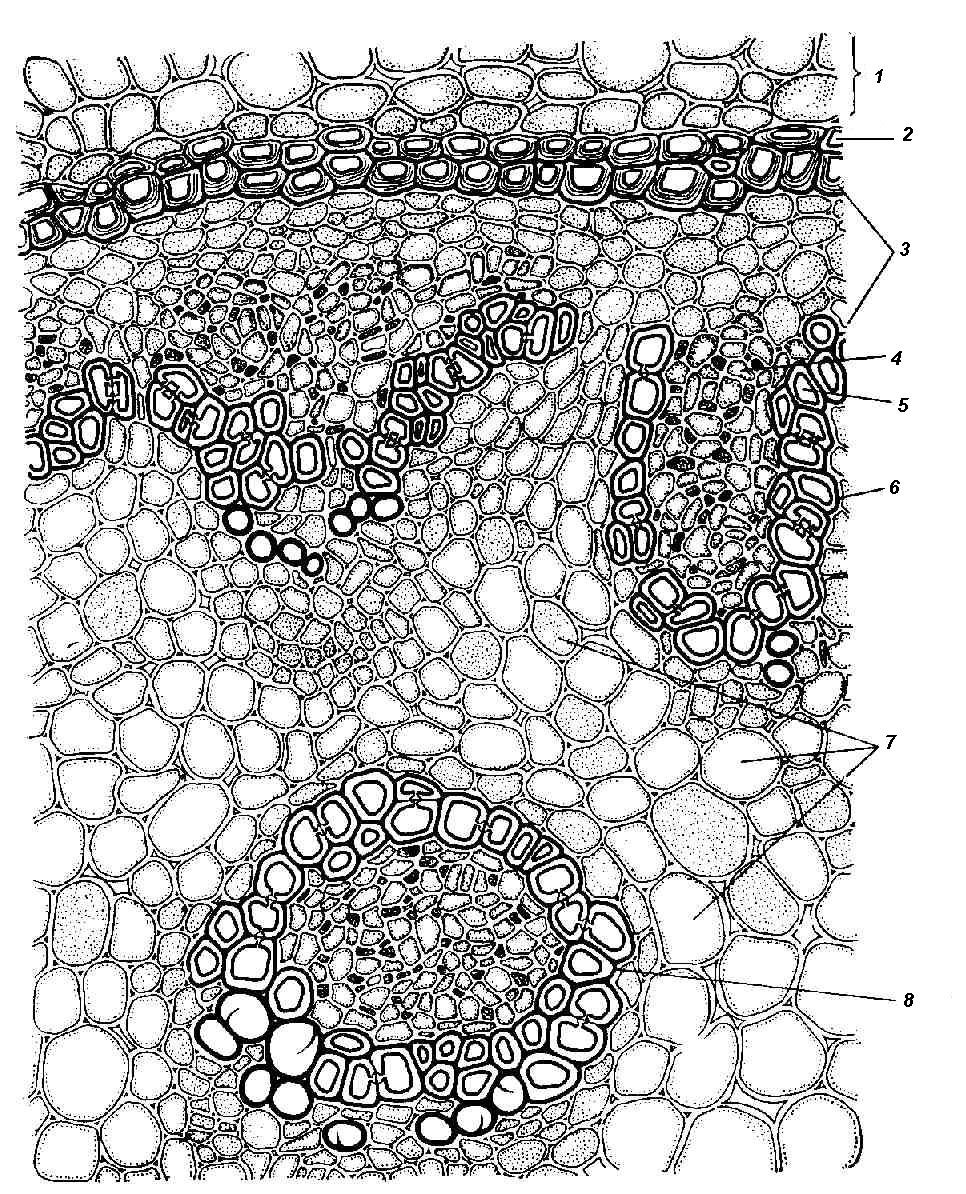 Рис. 4.48. Фрагмент поперечного среза корневища ландыша : 1 – паренхима первичной коры; 2 – эндодерма; 3 – перицикл; 4 – флоэма; 5 – ксилема; 6 – коллатеральный пучок; 7 – паренхима; 8 – концентрический амфивазальный пучок. Корневища двудольных растений могут иметь как пучковое (рис. 4.49), так и непучковое строение (рис. 4.50 ). Пучки открытые, как правило, коллатеральные, располагаются на поперечном срезе по окружности. 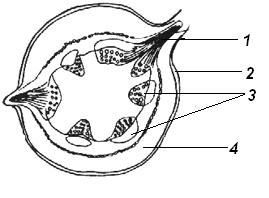 Рис. 4.49. Схема поперечного среза корневища валерианы : 1 – придаточный корень; 2 – перидерма; 3 – открытые коллатеральные пучки; 4 – первичная кора. 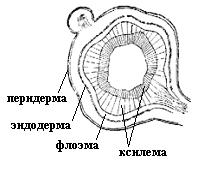 Рис. 4.50. Схема поперечного среза корневища синюхи (непучковое строение). Покровной тканью на молодых участках служит эпидерма, на старых участках корневищ эпидерма сменяется перидермой. От корня корневище всегда можно отличить по присутствию в центре основной ткани – сердцевины (центр корня занят ксилемой). |