|
1. Задачи,приводящие к понятию производной а о скорости движения материальной точки
25. Эффект Доплера и его использование в медико-биологических исследованиях
Эффектом Доплера называют изменение частоты волн, воспринимаемых наблюдателем, вследствие относительного движения источника волн и наблюдателя.
При одновременном сближении источника волн и наблюдателя воспринимается частота
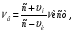
При одновременном удалении источника волн и наблюдателя воспринимается частота
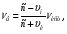 где c – скорость распространения УЗ волны, vн -скорость наблюдателя, vи -скорость источника, νи -частота излучения источника, νв -частота воспринимаемых волн. где c – скорость распространения УЗ волны, vн -скорость наблюдателя, vи -скорость источника, νи -частота излучения источника, νв -частота воспринимаемых волн.
Эффект Доплера используют для определения скорости кровотока
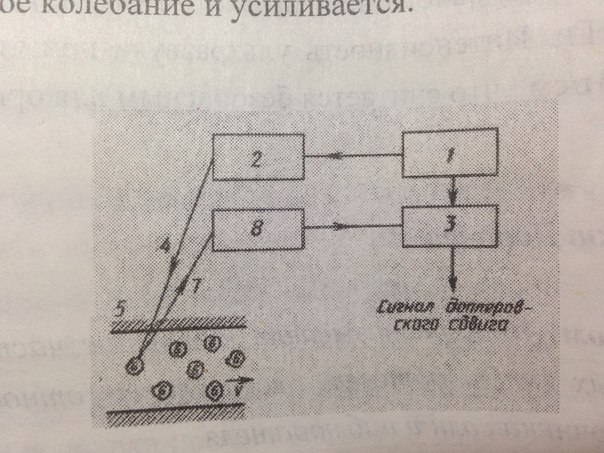
Используется система,содержащая генератор(1) электрических колебаний УЗ-частоты ,излучатель УЗ(2),устройство сравнения частот(3),УЗ_волна (4) проникает в кровеносный сосуд(5) и отражается от движущихся эритроцитов (6)Отраженная УЗ-волна (7) попадает в приемник(8),там преобразуется в электрическое колебание и усиливается
Если генератор излучает УЗ ,который падает на эритроциты ,то vг=vист=0,vприемн=vэр=v0
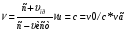 V= V=
(где v- частота волн,воспринимаемых эритроцитом )
Дальше рассмотрим волну,отраженную от эритроцита ,которфй станет теперь источником УЗ:
V”=_______ =________ ________ =_________
Где v”-частота,воспринимаемая приемником
Скорость кровотока равна:
Где vД- доплеровский сдвиг частоты
26. Законы отражения и преломления света. Явление полного внутреннего отражения. Предельный угол преломления. Предельный угол полного отражения.
Законы отражения и преломления света: 1)луч АО, падающий на преломляющую поверхность, нормаль к поверхности в точке падения РОР, луч отраженный ОВ и луч преломленный OD лежат в одной плоскости; 2) угол отражения РОВ численно равен углу падения РОА; 3) синус угла падения i относится к синусу угла преломления r, как скорость света к первой среде V1 относится к скорости света во второй среде V2: 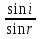 = =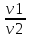 . Последний закон говорит о том, что свет распространяется в различных средах с различной скоростью. Предельный угол преломления: при переходе света из среды с меньшим показателем преломления в среду с большим показателем преломления угол падения луча больше угла преломления. Если луч падает на границу раздела сред под наибольшим возможным углом i= . Последний закон говорит о том, что свет распространяется в различных средах с различной скоростью. Предельный угол преломления: при переходе света из среды с меньшим показателем преломления в среду с большим показателем преломления угол падения луча больше угла преломления. Если луч падает на границу раздела сред под наибольшим возможным углом i=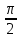 , то он будет преломляться под углом , то он будет преломляться под углом 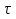 пр=. Этот угол является предельным углом преломления. Из закона преломления света следует: n21= пр=. Этот угол является предельным углом преломления. Из закона преломления света следует: n21=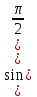 = =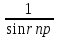 = =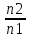 , откуда sin rпр= , откуда sin rпр=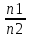 . Если свет переходит из оптически более плотной среды в оптически менее плотную, то угол преломления больше угла падения. При некотором угле падения i луча угол преломления равен , т.е. преломленный луч скользит вдоль границы раздела сред. При дальнейшем увеличении угла падения преломление не происходит, весь падающий свет отражается от границы раздела сред (полное отражение). Угол i называется предельным углом полного отражения и обозначается iпр. Так как n21= . Если свет переходит из оптически более плотной среды в оптически менее плотную, то угол преломления больше угла падения. При некотором угле падения i луча угол преломления равен , т.е. преломленный луч скользит вдоль границы раздела сред. При дальнейшем увеличении угла падения преломление не происходит, весь падающий свет отражается от границы раздела сред (полное отражение). Угол i называется предельным углом полного отражения и обозначается iпр. Так как n21=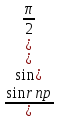 =, то sin iпр=. Таким образом, предельный угол преломления и предельный угол полного отражения для данных сред зависят от их показателей преломления. =, то sin iпр=. Таким образом, предельный угол преломления и предельный угол полного отражения для данных сред зависят от их показателей преломления.
27. Принцип действия рефрактометра. Ход лучей рефрактометра в проходящем и отраженном свете.
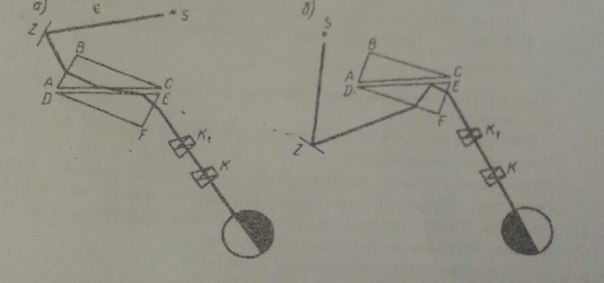 Принцип действия рефрактометра состоит в измерении предельного угла преломления на границе исследуемой жидкости и стеклянной призмы с известным коэффициентом преломления. При первом способе - в проходящем свете (рис.а) - пучок световых лучей, испускаемых источником света S, с помощью зеркальца Z направляется на грань АВ призмы ABC. Преломившись на грани АВ, лучи проходят в призму ABC и достигают грани АС. Но так как эта грань сделана матовой и поэтому вызывает рассеяние света, лучи войдут в жидкость и достигнут грани ДЕ под различными углами. Очевидно, что наибольший возможный угол падения для лучей, падающих на грань ДЕ, равен 90° . Эти скользящие вдоль поверхности ДЕ. Лучи после преломления определяют границу распространения света, так как им соответствует предельный угол преломления. При втором способе-в отраженном свете (рис. б) - пучок световых лучей, испускаемых источником S, с помощью зеркальца Z направляется на грань DF. Так как грань DF также матовая, то лучи входят в призму DEF под разными углами. В этом случае лучам, вошедшим в призму DEF и достигшим грани DE, приходится переходить из среды оптически более плотной (стекло) в среду оптически менее плотную (жидкость). Лучи, падающие на поверхность DE под углом меньше предельного, пройдут в жидкость и в призму ЛВС. Лучи, у которых угол падения больше предельного, претерпят полное внутреннее отражение. Лучи, направление которых соответствует величине предельного угла, и определяют границу раздела света и тени. В случае бесцветных и слабоокрашенных жидкостей удобно пользоваться первым способом. При измерении показателя преломления интенсивно окрашенных жидкостей, сильно поглощающих свет, лучше пользоваться вторым способом. Принцип действия рефрактометра состоит в измерении предельного угла преломления на границе исследуемой жидкости и стеклянной призмы с известным коэффициентом преломления. При первом способе - в проходящем свете (рис.а) - пучок световых лучей, испускаемых источником света S, с помощью зеркальца Z направляется на грань АВ призмы ABC. Преломившись на грани АВ, лучи проходят в призму ABC и достигают грани АС. Но так как эта грань сделана матовой и поэтому вызывает рассеяние света, лучи войдут в жидкость и достигнут грани ДЕ под различными углами. Очевидно, что наибольший возможный угол падения для лучей, падающих на грань ДЕ, равен 90° . Эти скользящие вдоль поверхности ДЕ. Лучи после преломления определяют границу распространения света, так как им соответствует предельный угол преломления. При втором способе-в отраженном свете (рис. б) - пучок световых лучей, испускаемых источником S, с помощью зеркальца Z направляется на грань DF. Так как грань DF также матовая, то лучи входят в призму DEF под разными углами. В этом случае лучам, вошедшим в призму DEF и достигшим грани DE, приходится переходить из среды оптически более плотной (стекло) в среду оптически менее плотную (жидкость). Лучи, падающие на поверхность DE под углом меньше предельного, пройдут в жидкость и в призму ЛВС. Лучи, у которых угол падения больше предельного, претерпят полное внутреннее отражение. Лучи, направление которых соответствует величине предельного угла, и определяют границу раздела света и тени. В случае бесцветных и слабоокрашенных жидкостей удобно пользоваться первым способом. При измерении показателя преломления интенсивно окрашенных жидкостей, сильно поглощающих свет, лучше пользоваться вторым способом.
28. Биологические мембраны, их структура и функции. Модели мембран.
Мембраны представляют собой плоские или изогнутые слои толщиной до 9 нм, образованные молекулами белков, жиров (липидов) и углеводов. Мембраны – это клеточные структуры, повсеместно встречающиеся в живых клетках и регулирующие обмен между клеткой и внешней средой (клеточные или плазматические мембраны), либо между различными частями клетки (внутриклеточные мембраны).
Основу мембраны образует двойной слой липидов. В этот слой встроены белковые молекулы, придающие специфические свойства различным участкам мембран, и тем самым, позволяющие последним принимать участие в разнообразных метаболитических процессах.
Молекулы липидов упакованы в слой так, что гидрофобные части (жировые хвосты) этих молекул отделены от воды, в то время как гидрофильные части (полярные головки) погружены в неё.
Двойной слой липидов как бы образует своеобразную двумерную жидкость с вязкостью, близкой к вязкости жидкого масла, поэтому молекулы липидов и белков легко перемещаются в плоскости слоя (латеральная диффузия).
При некоторых условиях (понижение температуры) в мембранах могут происходить процессы, сопровождающиеся изменением ориентации полярных головок и (или) затвердеванием углеводных хвостов липидов, что приводит к изменению функциональных свойств мембраны.Наиболее распространенной является жидкомозаичная модель мембраны: в липидном слое плавают более или менее погруженные белки.  1-поверхностные белки, 2- полупогруженные белки, 3- полностью погруженные белки, 4 – белки, формирующие «ионный канал» 5 1-поверхностные белки, 2- полупогруженные белки, 3- полностью погруженные белки, 4 – белки, формирующие «ионный канал» 5
Мембраны выполняют две важнейшие функции:
- матричную (являются матрицей, основой для удерживания белков, выполняющих различные функции);
- барьерную (защищают клетку и отдельные ее части от проникновения нежелательных частиц).
29. Перенос частиц через мембраны. Уравнение Фика. Применение уравнения Фика к биологической мембране. Уравнение Нернста-Планка.
Важной характеристикой мембран является их способность пропускать или не пропускать молекулы, атомы и ионы. Эти вопросы относятся к явлениям переноса.
Рассмотрим наиболее важные для биологических мембран явления: перенос вещества (диффузию) и перенос заряда (электропроводность).В биофизике также используется термин «транспорт частиц» Основное уравнение диффузии имеет вид:
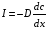
Это уравнение Фика :I- плотность потока частиц ,D- коэффицент диффузии D=___ __________,σ- среднее расстояние между молекулами, m-масса молекулы, n-концентрация молекул ,с=m*n-массовая концентрация, τ - среднее время оседлой жизни молекулы 
Применим уравнение Фика к биологической мембране.
Пусть концентрация частиц, диффундирующих через мембрану, изменяется в мембране линейно 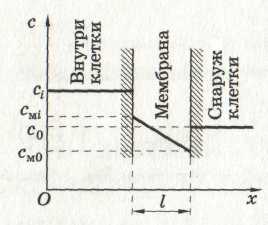
 Тогда : Тогда :
l - толщина мембраны, сi-концентрация частиц внутри клетки, с0- снаружи клетки, сmi - концентрация частиц на внутренней части мембраны, cmo- на наружной части. Отсюда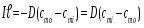
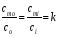 Практически легче определять концентрации частиц не внутри мембраны (cmiи cmo) , а вне мембраны: в клетке (сi) и снаружи клетки (co). Предположим, что Практически легче определять концентрации частиц не внутри мембраны (cmiи cmo) , а вне мембраны: в клетке (сi) и снаружи клетки (co). Предположим, что
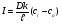 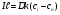 где k -коэффициент распределения частиц между мембраной и окружающей средой. Тогда cmo = kco, cmi = kci и имеем где k -коэффициент распределения частиц между мембраной и окружающей средой. Тогда cmo = kco, cmi = kci и имеем
отсюда:
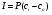 следовательно: следовательно:
P=Dk/l-- коэффициент проницаемости
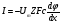 На мембране существует разностьпотенциалов, следовательно, в мембране имеется электрическое поле, которое влияет на диффузию заряженных частиц (ионов и электронов). Плотность потока заряда дается выражением На мембране существует разностьпотенциалов, следовательно, в мембране имеется электрическое поле, которое влияет на диффузию заряженных частиц (ионов и электронов). Плотность потока заряда дается выражением
где φ - потенциал поля, F = eNA-постоянная Фарадея, Z -валентность, Um- подвижность диффундирующих частиц для одного моля.
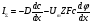 В общем случае перенос ионов зависит от неравномерности их распределения и воздействия электрического поля. Суммарная плотность потока частиц определяется уравнением Нернста-Планка В общем случае перенос ионов зависит от неравномерности их распределения и воздействия электрического поля. Суммарная плотность потока частиц определяется уравнением Нернста-Планка
30. Пассивный транспорт и его основные виды. Понятие об активном транспорте.
Различают два вида транспорта веществ через мембраны.
Пассивный транспорт имеет следующие разновидности:
1. Простая диффузия;
2. Диффузия через канал;
3. Облегчённая диффузия (осуществляется молекулами-переносчиками).
Пассивный транспорт не связан с затратой химической энергии, он происходит в результате перемещения частиц в сторону меньшего электрохимического потенциала.
Простая диффузиячерез липидный слой подчиняется в общем случае уравнению Нернста-Планка. Она происходит медленно и не может снабдить клетки в нужном количестве питательными веществами.
Перенос вещества через каналы происходит без затраты энергии и направлен в сторону уменьшения концентрации молекул. Каналы, образованные интегральными белками, проявляют избирательность к разным ионам. Такая селективность канала к различным ионам определяется его формой и размерами, а также электростатическими свойствами аминокислот, выстилающих поверхность канала.
Процесс облегченной диффузии также происходит без затраты энергии, направлен в сторону уменьшения концентрации молекул и протекает следующим образом: молекула переносимого вещества связывается со специальной молекулой-переносчиком в единый комплекс, который легко проходит через мембрану, а на другой ее стороне распадается, отщепляя переносимую молекулу.
При активном транспорте перенос вещества происходит в сторону больше концентрации. Этот процесс не является диффузией и протекает за счёт затраты энергии. Самой распространённой системой активного транспорта является натрий-калиевый насос. Натрий-калиевые насосы работают за счёт энергии гидролиза молекул АТФ с образованием молекул АДФ и неорганического фосфора Ф:
АТФ = АДФ + Ф.
Натрий-калиевый насос работает обратимо:крадиенты концентрации ионов К+ и Na- способствуют синтезу молекул АТФ :
АДФ+Ф=АТФ
31. Биоэлектрические потенциалы. Потенциал покоя. Механизм генерации потенциала действия.
Концентрации различных ионов внутри и снаружи клетки неодинаковы. Во внеклеточном пространстве имеется избыток ионов Na+ , Cl-, внутри клетки ионов К+. Они могут диффундировать через пористую белковую структуру мембраны. Скорости диффузии разных ионов различны. Помимо указанных ионов внутриклеточные и внеклеточные жидкости содержат большое количество отрицательных ионов (ионов фосфатов, карбонатов и т.д.). Но размеры этих ионов больше, чем ионные каналы и их диффузионным эффектом можно пренебречь.
Рассмотрим диффузионные процессы на примере ионов К+. Их концентрация внутри клетки в 30 раз больше, чем во внеклеточном пространстве.
Если бы мембрана была пассивной структурой ,т.е., разделяла бы два участка с одинаковым электрическим потенциалом, то ионы К+ диффундировали бы в обоих направлениях с одинаковой скоростью, и градиента концентрации не возникало бы. Но на поверхностях мембраны имеется двойной слой зарядов, и существует разность потенциалов, которая приблизительно равна-70 мВ. Эта разность потенциалов между цитоплазмой и окружающей средой называется потенциалом покоя. Он обусловливает наличие градиента концентрации различных ионов. Отметим, что жидкость с каждой стороны мембраны электрически нейтральна. Заряды имеются только на внутренней и внешней поверхности мембраны.
 Разность потенциалов, которая обеспечивает равновесное отношение концентраций, дается уравнением Нернста: Разность потенциалов, которая обеспечивает равновесное отношение концентраций, дается уравнением Нернста:
где сi - концентрация ионов внутри клетки, co - снаружи клетки, а φ - в мВ. Для отрицательных ионов знак разности потенциалов меняется на противоположный.
Полная плотность потока ионов K+, Na+, Cl- с учетом знаков равна
I = IK + INa - ICl
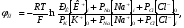 В стационарном состоянии I=0, т.е. число ионов, выходящих из клетки в единицу времени равно числу ионов, входящих в клетку. В общем случае потенциал покоя дается уравнением Гольдмана-Ходжкина-Катца В стационарном состоянии I=0, т.е. число ионов, выходящих из клетки в единицу времени равно числу ионов, входящих в клетку. В общем случае потенциал покоя дается уравнением Гольдмана-Ходжкина-Катца
где квадратными скобками [ ]i и [ ]o обозначены концентрации ионов соответственно внутри и вне клетки.
Потенциал покоя – стационарная разность электрических потенциалов между внутренней и наружной поверхностями мембраны в невозбужденном состоянии.
Если вычислить потенциал Нернста, то для ионов Cl- имеем φ (Cl-) = -70 мВ. Потенциал покоя также равен - 70 мВ т.е. ионы будут диффундировать во внутрь клетки и наружу с одинаковыми скоростями, а разность концентраций будет поддерживаться за счет разности потенциалов.
Если бы диффузия была единственным процессом, управляющим движением ионов через мембрану, то через некоторое время установилось бы равновесное отношение концентраций. Но помимо диффузии действуют также системы активного транспорта – натрий-калиевые насосы, и их действие противоположно направлению нормальной диффузии.
При возбуждении клетки разность потенциалов между клеткой и окружающей средой изменяется – возникает потенциал действия.
Потенциалом действия называется электрический импульс, возникающий между внутренней и наружной сторонами мембраны и обусловленный изменением ионной проницаемости мембраны.
Рассмотрим распространение потенциала действия в нервном волокне. Нервная клетка или нейрон состоит из клеточного тела и выроста, называемого аксоном. Нейрон можно привести в возбужденное состояние в любой точке аксона электрическим, химическим или механическим способом.
.Генерация и распространение потенциаВ состоянии покоя мембранный потенциал равен -70 мВ
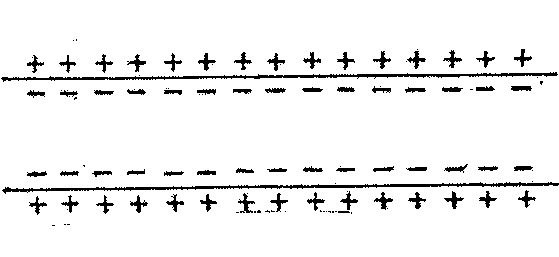 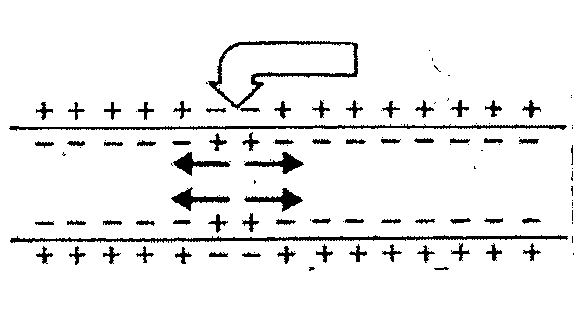
1Состояние покоя 2 началась деполяризация
 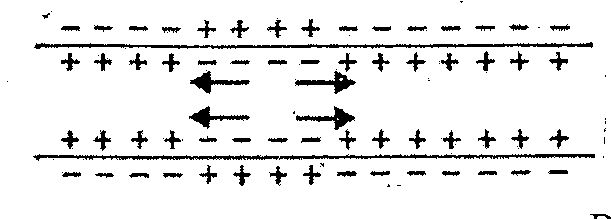
3Участок полностью деполяризован 4началась реполяризация
При возбуждении аксона внезапно увеличивается проницаемость ионов Na+ (в 5000 раз и более). Большое количество ионов натрия устремляется в клетку, перекрывая ее отрицательный потенциал. За доли секунды возникает локальный положительный потенциал. Этот положительный потенциал, равный +40 мВ называется потенциалом действия. Потенциал действия начинает распространяться по обе стороны от возбужденной точки, пока не деполяризуется весь участок. После этого увеличивается проницаемость ионов К+ и они начинают диффундировать из клетки во внеклеточное пространство. Начинается фаза реполяризации и значение потенциала действия становится отрицательным, меньшим чем потенциала покоя
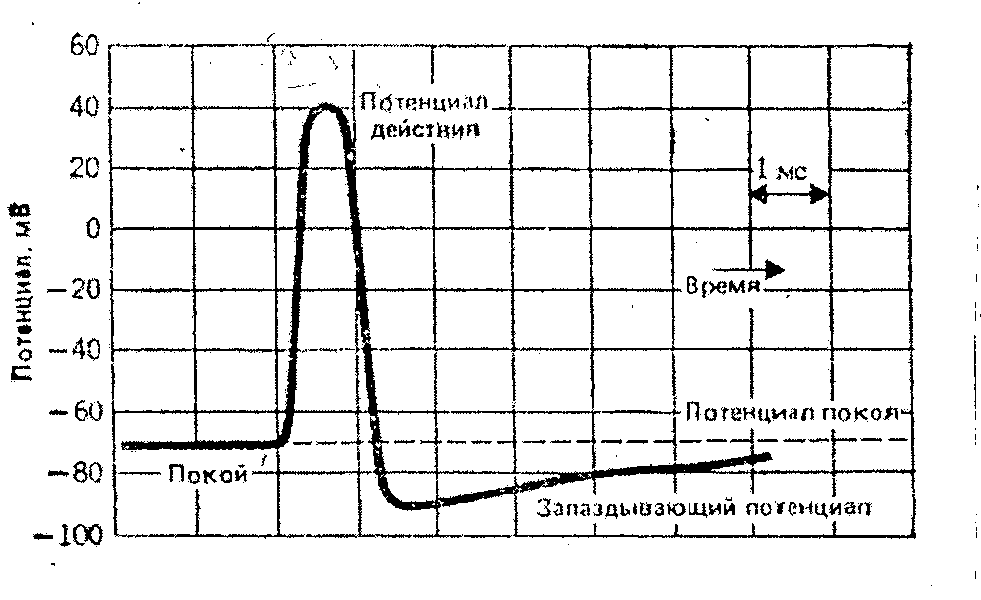
В этот момент начинают работать натрий-калиевые насосы, которые восстанавливают значение потенциала покоя, после чего весь процесс повторяется. Весь процесс занимает
10 мс. Отметим, что генерация потенциала действия происходит по принципу «все или ничего» – процесс либо вообще не начинается, либо идет до конца.
|
|
|
 Скачать 1.34 Mb.
Скачать 1.34 Mb.