Коллок№1Шиза. 12. 1 Раздражимость и возбудимость как основа реакции ткани на раздражение. Понятие о раздражителе, виды раздражителей, характеристика. Понятие порога раздражения
 Скачать 105.49 Kb. Скачать 105.49 Kb.
|

|
12.1 Раздражимость и возбудимость как основа реакции ткани на раздражение. Понятие о раздражителе, виды раздражителей, характеристика. Понятие порога раздражения. Раздражимость – это способность ткани изменять свой обмен веществ и энергии под действием раздражителей. Раздражимость это свойство характерное для всех тканей организма. Возбудимость – это способность возбудимых тканей на действие раздражителя отвечать возбуждением, которое проявляется в виде биоэлектрического процесса и специфической ответной реакции. Раздражитель – любой сдвиг, изменения во внешней или внутренней среде биосистемы достаточной силы, чтобы вызвать изменение её состояния. Виды раздражителей : 1) адекватные 2) неадекватные Адекватным раздражителем называют раздражитель, к восприятию которого биосистема приспособлена. Он способен вызвать реакцию биосистемы при минимальных значениях. Неадекватный раздражитель способен вызвать реакцию биосистемы, но при значительных абсолютных величинах. Также классифицируют на: 1) пороговые 2) подпороговые 3) надпороговые Пороговым раздражителем называют минимальную силу раздражителя, способную вызвать регистрируемую реакцию Подпороговым раздражителем никогда не вызывают регистрируемую реакцию Надпороговым – всегда вызывают регестрируемую реакцию. Порогом раздражения называют минимальную силу раздражителя, способную вызвать регистрируемую реакцию. 13.2 Законы раздражения возбудимых тканей: значение силы раздражителя, частоты раздражителя, его длительности, крутизны его нарастания. 1. Постоянный ток раздражает только при своем изменении 2. Раздражающие действия постоянного тока тем сильнее, чем быстрее он меняется во времени(чем выше скорость нарастания), значит макс. эффективность у мгновенного тока (Р-импульсы) 3. Катод повышает возбудимость и возбуждает, анод снижает возбудимость и уменьшает текущее возбуждение(тормозит). Закон полярного действия постоянного тока ( 1 закон Пфлюгера) – закон о месте и времени возникновения возбуждения в клетке: При действии на ткань постоянного тока возбуждение под катодом возникает в момент замыкания цепи, а под анодом в момент размыкания. Раздражающее действие катода выражено сильнее, чем анода. 2. Закон физиологического электротона (2 закон Пфлюгера) – закон об изменении физиологических свойств тканей при прохождении через них постоянного тока: При прохождении через ткань постоянного тока в области катода возбудимость и проводимость повышаются, а лабильность снижается(физиологический Кат-электротон), в области анода возбудимость и проводимость понижается, а лабильность повышается – физиологический АН-электротон. Дополнение Вериго – при прохождении через ткань сильного или длительного постоянного тока повышенная возбудимость в области катода сменяется пониженной – катодическая депрессия, а пониженная возбудимость в области анода сменяется повышенной – анодическая экзальтация. ИТОГ: Возбудимость клетки не зависит ни от величины мембранного потенциала, ни от величины критического уровня деполяризации, а определяется их соотношением: чем меньше между ними разница, тем выше возбудимость и наоборот Закон Дюбуа-Раймона: Раздражающее действие постоянного тока не зависит ни от силы тока (его плотности), ни от длительности действия(времени), а определяется скоростью его нарастания: чем быстрее меняется ток во времени, тем большей раздражающей силой он обладает. Закон «Кривая Сила-Время»: Раздражающее действие постоянного тока тем сильнее, чем больше время его действия Хроноксия – время, в теч которого на ткань должен действовать постоянный ток, равным 2 реобазам, чтобы вызвать возбуждение. 14.3 Современные представления о строении и функции мембран. Ионные каналы мембран. Ионные градиенты клетки, механизмы из возникновения. Функции: 1. Барьерная – мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. 2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»). 3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах). 4.Высвобождение нейромедиаторов в синаптических окончаниях. Жидкостно-мозаичная модель Сингера и Николсона: В фосфолипидном бислое интегрированы глобулярные белки, полярные участки которых образуют гидрофильную поверхность в водной фазе. Эти интегрированные белки выполняют различные функции, в том числе рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул. Некоторые белковые молекулы свободно диффундируют в плоскости липидного слоя; в обычном состоянии части белковых молекул, выходящие по разные стороны клеточной мембраны, не изменяют своего положения. Особая морфология клеточных мембран определяет их электрические характеристики, среди которых наиболее важными являются емкость и проводимость. Емкостные свойства в основном определяются фосфолипидным бислоем, который непроницаем для гидратированных ионов и в то же время достаточно тонок (около 5 нм), чтобы обеспечивать эффективное разделение и накопление зарядов и электростатическое взаимодействие катионов и анионов Проводимость (g) — величина, обратная электрическому сопротивлению и равная отношению величины общего трансмембранного тока для данного иона к величине, обусловившей его трансмембранной разности потенциалов. Строение и функции ионных каналов. Ионы Na+, K+, Са2+, Сl- проникают внутрь клетки и выходят наружу через специальные, заполненные жидкостью каналы. Размер каналов довольно мал (диаметр 0,5—0,7 нм). Рассмотрим принцип работы ионных каналов на примере натриевого канала. Полагают, что в состоянии покоя натриевый канал закрыт. При деполяризации клеточной мембраны до определенного уровня происходит открытие m-активационных ворот (активация) и усиление поступления ионов Na+ внутрь клетки. Через несколько миллисекунд после открытия m-ворот происходит закрытие п-ворот, расположенных у выхода натриевых каналов (инактивация) (рис. 2.4). Инактивация развивается в клеточной мембране очень быстро и степень инактивации зависит от величины и времени действия деполяризующего стимула. При генерации одиночного потенциала действия в толстом нервном волокне изменение концентрации ионов Na+ во внутренней среде составляет всего 1/100000 от внутреннего содержания ионов Na гигантского аксона кальмара. Однако для тонких нервных волокон это изменение концентрации может быть весьма существенным. Кроме натриевых, в клеточных мембранах установлены другие виды каналов, избирательно проницаемых для отдельных ионов: К+, Са2+ Ходжкин и Хаксли сформулировали принцип «независимости» каналов, согласно которому потоки натрия и калия через мембрану независимы друг от друга. Свойство проводимости различных каналов неодинаково. В частности, для калиевых каналов процесс инактивации, как для натриевых каналов, не существует. Имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-кальцийзависимых каналов ускоряет реполяризацию, тем самым восстанавливая исходное значение потенциала покоя. Особый интерес представляют кальциевые каналы. Входящий кальциевый ток, как правило, недостаточно велик, чтобы нормально деполяризовать клеточную мембрану. Чаще всего поступающий в клетку кальций выступает в роли вторичного посредника. Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, например входящим натриевым током. Процесс инактивации кальциевых каналов достаточно сложен. С одной стороны, повышение внутриклеточной концентрации свободного кальция приводит к инактивации кальциевых каналов. С другой стороны, белки цитоплазмы клеток связывают кальций, что позволяет поддерживать длительное время стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется. Кальциевые каналы играют существенную роль в клетках сердца. 15.4 Мембранный потенциал, теория его происхождения. Мембранный потенциал - это разность потенциалов между наружной и внутренней поверхностями элементарной пограничной мембраны клетки Мембранный потенциал – сила электростатического взаимодействия, которая уравновешивает электрические токи, протекающие через мембрану изнутри кнаружи, снаружи – внутрь. Современная теория Ходжкина-Хаксли-Катца: 1. В основе возникновения биопотенциала является обмен веществ и энергии между клеткой и окружающей средой 2. Местом возникновения биопотенциала является поверхностная мембрана клетки 3. Непосредственной причиной возникновения биопотенциала является разность концентрации ионов (ионная асимметрия вне и внутри клетки) 4. Поверхностная пограничная мембрана клетки обладает свойством полупроницаемости, которое может меняться при различных состояниях. 16.5. Потенциал действия, его фазы. Динамика проницаемости мембраны в различные фазы потенциала действия. Под потенциалом действия понимают быстрое колебание потенциала, сопровождающееся, как правило, перезарядкой мембраны. Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны. При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Компоненты потенциала действия: 1) локальный ответ; 2) высоковольтный пиковый потенциал (спайк); 3) следовые колебания. Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку. Высоковольтный пиковый потенциал (спайк). Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз: 1) восходящей части – фазы деполяризации; 2) нисходящей части – фазы реполяризации. Быстрое и большое количество поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K. Полного восстановления мембранного потенциала не происходит. В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный. 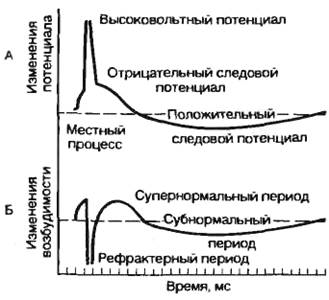 17.6 Возбудимость, методы её оценки. Изменения возбудимости при действии постоянного тока (электротон, катодическая депрессия, аккомодация). Возбудимость – это способность возбудимых тканей на действие раздражителя отвечать возбуждением, которое проявляется в виде биоэлектрического процесса и специфической ответной реакции. Подпороговое пассивное поведение клеточной мембраны называется электротоническим, или электротоном. Закон физиологического электротона (2 закон Пфлюгера) – закон об изменении физиологических свойств тканей при прохождении через них постоянного тока: При прохождении через ткань постоянного тока в области катода возбудимость и проводимость повышаются, а лабильность снижается(физиологический Кат-электротон), в области анода возбудимость и проводимость понижается, а лабильность повышается – физиологический АН-электротон. Дополнение Вериго – при прохождении через ткань сильного или длительного постоянного тока повышенная возбудимость в области катода сменяется пониженной – катодическая депрессия, а пониженная возбудимость в области анода сменяется повышенной – анодическая экзальтация. ИТОГ: Возбудимость клетки не зависит ни от величины мембранного потенциала, ни от величины критического уровня деполяризации, а определяется их соотношением: чем меньше между ними разница, тем выше возбудимость и наоборот Закон Дюбуа-Раймона: Раздражающее действие постоянного тока не зависит ни от силы тока (его плотности), ни от длительности действия(времени), а определяется скоростью его нарастания: чем быстрее меняется ток во времени, тем большей раздражающей силой он обладает. Аккомодация ткани – явление приспособления (снижения возбудимости) ткани к действию медленно-нарастающего раздражителя. 18.7 Соотношения фаз изменения возбудимости при возбуждении с фазами потенциала действия. 1) местный ответ - физиологический катэлектротон. 2) высоковольтный пик - катодическая депрессия 3) следовая деполяризация - катэлектротон 4) следовая гиперполяризация - анэлектротон Когда ткань возбуждается генерирует ПД, то временно в ней меняется возбудимость: сначала ткань становится совершенно невозбудимой (абсолютная рефрактерность) любой по силе стимул не способен вызывать в ней новый приступ возбуждения, эта фаза обычно наблюдается во время пика ПД, затем происходит постепенное восстановление возбудимости до исходного состояния (фаза относительной рефрактерности) – в этот момент раздражитель может вызвать возбуждение (генерацию нового ПД), но для этого он должен быть выше порогового (исходного). Затем в фазу следовой негативности возбудимость повышается (супервозбудимость или фаза экзальтации). В этот момент подпороговые раздражители могут вызвать возбуждение. В тканях, в которых ярко проявилась следовая гиперполяризация наблюдается еще одна фаза- субнормальности (фаза сниженной возбудимости). 19.8 Строение и классификация синапсов. Механизм передачи сигналов в синапсах ( электрических и химических) Ионные механизмы постсинаптических потенциалов, их виды. Синапсом называют образование, обеспечивающее передачу возбуждения от одной структуры к другой. Синапсы по способу передачи сигнала: I.Химические (передача сигналов с помощью медиатора) II.Электрические (нексусы) – нет у высш животных и у человека Синапсы: I. Центральные (>60) II. Периферические : 1) Соматические (нервно-мышечные) 2) Вегетативные: 1. Ганглионарные 2. Эффекторные(симпатические и парасимпатические) Синапсы по характеру действия на воспринимающую структуру: I. возбуждающие II.тормозящие По местоположению выделяют нервно-мышечные синапсы и нейронейрональные, последние в свою очередь делятся на аксосоматические, аксоаксональные, аксодендритические, дендросоматические. В химическом синапсе выделяют 3 элемента : пресинаптическую область (пресинаптическая терминаль); синаптическую щель; постсинаптическую область. Пресинаптическая область представляет собой демиелинизированный терминальный участок отростка нервной клетки. По форме пресинаптическая терминаль напоминает собой колбу, прилежащую основанием к участку мембраны возбудимой клетки. Наиболее существенной чертой пресинаптической области является скопление пресинаптических пузырьков диаметром 50 мм (везикул), в которых содержится медиатор (химическое соединение, которое является материальным носителем сигнала к реципиентской клетке). Кроме везикул с медиатора в пресинаптической области содержится большое количество митохондрий и лизосом, что свидетельствует о высокой активности обменных процессов в этой области. Кроме того, в этой области обнаружены предшественники медиаторов и продукты их метаболизма. Синаптическая щель. В химических синапсах составляет от 20 до 50 мм. Здесь содержаться вода, электролиты, олигосахариды, ферменты, участвующие в расщеплении медиатора. Постсинаптическая область. Включает субсинаптическую мембрану (участок постсинаптической мембраны, имеющий специальный аппарат -рецепторы, характеризующиеся сродством к медиатору. В этой же области имеются химически-чувствительные ионные каналы. Собственно постсинап-тическая мембрана - участок постсинаптической мембраны, которая содержит потенциал-зависимые ионные каналы и на которой происходит генерация постсинаптических потенциалов. Современные представления о механизмах передачи возбуждения в синапсе. ПД распространяется по нервному волокну к пресинаптической области. Изменение проницаемости мембраны пресинаптического образования к ионам Са++ и поступление Са++ в пресинаптическом образовании. Движение везикул с активным медиатором пресинаптической области к пресинаптической мембране и выделение медиатора в синаптическую щель методом экзоцитоза. Движение медиатора к субсинаптической мембране постсинаптической области и взаимодействие с соответствующими рецепторами мембраны. Далее возможны два варианта: 1- либо само взаимодействие медиатора с рецептором приводит к активации ионных каналов субсинаптической мембраны (это имеет место в некоторых холинэргических синапсах; 2 - либо вначале активируется вторичный посредник, а затем уже включается цепь биохимических процессов, приводящая к изменению проницаемости ионных каналов. Изменение проницаемости ионных каналов приводит к формированию местных токов и генерации на постсинаптической мембране постсинаптического потенциала. В зависимости от того по отношению к какому иону меняется проницаемость мембран, возможны два варианта постсинаптических потенциалов - возбуждающий, деполяризационный (ВПСП) и тормозной, гиперполяризационный (ТПСП). В основе ВПСП лежит повышение проницаемости мембраны по отношению к ионам Nа+, а ТПСП- по отношению к ионам К+ и Cl- . Характер активности тех или иных каналов определяется химической структурой медиатора, особенностью рецепторного образования, а также связанного с ним вторичного посредника. Электрические синапсы обладают односторонним проведением возбуждения. Это легко доказать при регистрировании электриче ского потенциала на синапсе: при раздражении афферентных путей мембрана синапса деполяризуется, а при раздражении эфферентных волокон — гиперполяризуется. Оказалось, что синапсы нейронов с одинаковой функцией обладают двусторонним проведением возбуж дения (например, синапсы между двумя чувствительными клетка ми), а синапсы между разнофункциональными нейронами (сенсор ные и моторные) обладают односторонним проведением. Функции электрических синапсов заключаются прежде всего в обеспечении срочных реакций организма. Этим, видимо, объясняется располо жение их у животных в структурах, обеспечивающих реакцию бегства, спасения от опасности и т. д. Электрический синапс сравнительно мало утомляем, устойчив к изменениям внешней и внутренней среды. Видимо, эти качества наряду с быстродействием обеспечивают высокую надежность его работы. 20.10 Определение медиаторов и синоптических рецепторов, их классификация и роль в проведении сигналов в возбуждающих и тормозных синапсах. Рецептором называют генетически детерменированные макромолекулы, которые распологаются на субсинаптической мембране и взаимодействуют с молекулами медиатора как ключ с замком, запуская при этом реактивную систему синапса. Медиатор – это группа химических веществ, которая принимает участие в передаче возбуждения или торможения в химических синапсах с пресинаптической на постсинаптическую мембрану. 1. Возбуждающие медиаторы, вызывающие деполяризацию постсинаптической мембраны и образование возбуждающего постсинаптического потенциала: 1) АХ; 2) глютаминовая кислота; 3) аспарагиновая кислота. 2. Тормозящие медиаторы, вызывающие гиперполяризацию постсинаптической мембраны, после чего возникает тормозной постсинаптический потенциал, который генерирует процесс торможения: 1) ГАМК; 2) глицин; Рецепторы, располагающиеся на наружных образованиях тела, называются экстерорецепторами, а в различных внутренних органах — интерорецепторами (висцерорецепторами). В зависимости от природы действу ющего раздражителя рецепторы подразделяются на: механорецепторы (воспринимают воздействие при механическом сме щении или деформации рецепторного участка); хеморецепторы (активируются различными химическими соединениями); фоторецепторы (воспринимают электромагнитное излучение в диапа зоне волн от 400 до 750 нм); терморецепторы и др. 21Определение медиаторов и синаптическихрецепторов,их классификация и роль в проведение сигналов в возбуждающих и тормозных синапсов. Медиатор – это группа химических веществ, которая принимает участие в передаче возбуждения или торможения в химических синапсах с пресинаптической на постсинаптическую мембрану. Классификация медиаторов:1) химическая, основанная на структуре медиатора;2) функциональная, основанная на функции медиатора. Химическая классификация.1. Сложные эфиры – ацетилхолин (АХ).2. Биогенные амины(катехоламины ,серотонин).3. Аминокислоты(гаммааминомасляная кислота (ГАМК);глютаминовая кислота; глицин;аргинин.)4. Пептиды(опиоидные пептиды,вещество «P»;вазоактивныйинтестинальныйпептид;соматостатин.)5. Пуриновые соединения: АТФ.6. Вещества с минимальной молекулярной массой( NO;CO.) Функциональная классификация.1. Возбуждающие медиаторы, вызывающие деполяризацию постсинаптической мембраны и образование возбуждающего постсинаптического потенциала:АХ; глютаминовая кислота;аспарагиновая кислота.2. Тормозящие медиаторы, вызывающие гиперполяризацию постсинаптической мембраны, после чего возникает тормозной постсинаптический потенциал, который генерирует процесс торможения: ГАМК;глицин; вещество «P»;дофамин;серотонин;АТФ. Норадреналин, изонорадреналин, адреналин, гистамин являются как тормозными, так и возбуждающими.АХ (ацетилхолин) является самым распространенным медиатором в ЦНС и в периферической нервной системе. Содержание АХ в различных структурах нервной системы неодинаково. С филогенетической точки зрения в более древних структурах нервной системы концентрация ацетилхолина выше, чем в молодых. АХ находится в тканях в двух состояниях: связан с белками или находится в свободном состоянии (активный медиатор находится только в этом состоянии). Синапс – это структурно-функциональное образование, обеспечивающее переход возбуждения или торможения с окончания нервного волокна на иннервирующую клетку. 1. По локализации:Центральные синапсы лежат в пределах центральной нервной системы, а также находятся в ганглиях вегетативной нервной системы. Центральные синапсы – это контакты между двумя нервными клетками, причем эти контакты неоднородны и в зависимости от того, на какой структуре первый нейрон образует синапс со вторым нейроном, различают:1) аксосоматический, образованный аксоном одного нейрона и телом другого нейрона;2) аксодендритный, образованный аксоном одного нейрона и дендритом другого;3) аксоаксональный (аксон первого нейрона образует синапс на аксоне второго нейрона);4) дендродентритный (дендрит первого нейрона образует синапс на дендрите второго нейрона). Различают несколько видов периферических синапсов:1) мионевральный (нервно-мышечный), образованный аксоном мотонейрона и мышечной клеткой;2) нервно-эпителиальный, образованный аксоном нейрона и секреторной клеткой. 2. Функциональная классификация синапсов:возбуждающиесинапсы;тормозящие синапсы. 3. По механизмам передачи возбуждения в синапсах:химические; электрические. Особенность химических синапсов заключается в том, что передача возбуждения осуществляется при помощи особой группы химических веществ – медиаторов. Синапсы имеют ряд физиологических свойств: 1) клапанное свойство синапсов, т. е. способность передавать возбуждение только в одном направлении с пресинаптической мембраны на постсинаптическую; 2) свойство синаптической задержки, связанное с тем, что скорость передачи возбуждения снижается; 3) свойство потенциации (каждый последующий импульс будет проводиться с меньшей постсинаптической задержкой). Это связано с тем, что на пресинаптической и постсинаптической мембране остается медиатор от проведения предыдущего импульса; 4) низкая лабильность синапса (100–150 имульсов в секунду). 22.11 Физические и физиологические свойства мышц. Типы мышечных сокращений. Сила и работа мышц. Закон силы. Свойства скелетных мышц : 1) обеспечивают определенную позу тела человека; 2) перемещают тело в пространстве; 3) перемещают отдельные части тела относительно друг друга; 4) являются источником тепла, выполняя терморегуляционную функцию. 5) возбудимостью — способностью отвечать на действие раздражителя изменением ионной проводимости и мембранного потенциала. 6) проводимостью — способностью проводить потенциал дейст вия вдоль и в глубь мышечного волокна по Т-системе; 7) сократимостью — способностью укорачиваться или разви вать напряжение при возбуждении; 8) эластичностью — способностью развивать напряжение при растягивании. Свойства гладких мышц : 1) Статическая ( сохранительная, тоническая) 2) Низкая электропроводимость и высокая хемовозбудимость 3) Автоматия 3) Низкая лабильность 4) Низкая проводимость 5) Высокая пластичность 6) Слабые, практически неутомлямые Типы мышечных сокращений. Различают изотонический, изометрический и смешанный режимы сокращения мышц. При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, когда мышца не перемещает груз. В естественных условиях близкими к изотоническому типу сокращений являются сокращения мышц языка.(один фиксирован, другой висит свободно) При изометрическом сокращении длина мышечных волокон остается постоянной, меняется напряжение мышцы. Такое сокращение мышцы можно получить при попытке поднять непосильный груз.(оба конца мышцы фиксированы и она не может укоротиться) Сила - мера механического воздействия на мышцу со стороны других тел, которая выражается в ньютонах или кг-силах. При изотоническом сокращении в эксперименте сила определяется массой максимального груза, который мышца может поднять (динамическая сила), при изометрическом - максимальным напряжением, которое она может развить (статическая сила). Одиночное мышечное волокно развивает напряжение в 100-200 кг-сил во время сокращения. Изометрически сокращающаяся мышца развивает максимально возможное для нее напряжение в результате активации всех мышечных волокон. Такое напряжение мышцы называют максимальной силой. Отношение максимальной силы мышцы к ее анатомическому поперечнику называется относительной силой мышцы, измеряемой в кг/см2. Работа есть энергия, затрачиваемая на перемещение тела с определенной силой на определенное рас стояние: А = FS. При сокращении скелетной мускулатуры в естественных условиях преимущественно в режиме изометрического сокращения, например при фиксированной позе, говорят о статической работе, при со вершении движений — о динамической. Сила сокращения и работа, совершаемая мышцей в единицу вре мени (мощность), не остаются постоянными при статической и дина мической работе. В результате продолжительной деятельности рабо тоспособность скелетной мускулатуры понижается. Это явление назы вается утомлением. При этом снижается сила сокращений, увеличиваются латентный период сокращения и период расслабления. Закон Силы: в определенных пределах амплитуда сокращения скелетной мышцы тем больше, чем больше сила сокращения. 23.12 Одиночное сокращение и его фазы . Тетанус, факторы, влияющие на его величину. Понятие оптимума и пессимума. Раздражение мышечного волокна одиночным пороговым или сверхпороговым стимулом приводит к возникновению одиночного со кращения. Периоды : Пер вый — латентный период представляет собой сумму временных задержек, обусловленных возбуждением мембраны мышечного волок на, распространением ПД по Т-системе внутрь волокна, образованием инозитолтрифосфата, повышением концентрации внутриклеточного кальция и активации поперечных мостиков. Для портняжной мышцы лягушки латентный период составляет около 2 мс. Второй — период укорочения, или развития напря жения. В случае свободного укорочения мышечного волокна говорят об изотоническом режиме сокращения, при котором напряжение практически не изменяется, а меняется только длина мышечного во локна. Если мышечное волокно закреплено с двух сторон и не может свободно укорачиваться, то говорят об изометрическом режиме со кращение. Третий — период расслабления, когда уменьшается кон центрация ионов Са2+ и отсоединяются головки миозина от актиновых филаментов. Тетанусом наз. Стойкое длительное укорочение мышцы в ответ на ряд раздражителей. Зубчатый возникает тогда, когда интервал между раздражителями меньше чем продолжительность одиночного сокращения и больше чем период укорочения. Гладкий возникает когда интервал между раздражителями меньше чем период укорочения мышцы. Оптимум(наилучший) – максимальная ответная реакция биосистемы в ряду возрастающих по силе или частоте раздражений. Пессимум (наихудший) – ответная реакция биосистемы на сверхоптимальный раздражитель 24.13 Двигательные единицы, их классификация. Роль в формировании динамических и статических сокращений скелетных мышц в естественных условиях. Моторной (двигательной) единицей называют совокупность нейрона, аксона и мышечных элементов, которые они иннервируют. Двигательные единицы млекопитающих можно разделить на три группы (Burke, 1981): медленно сокращающиеся невосприимчивые к утомлению (тип S); быстро сокращающиеся невосприимчивые к утомлению (тип FR) и быстро сокращающиеся восприимчивые к утомлению (тип FF). Число мышечных волокон, входящих в состав двигательной единицы, различно и зависит от функции, которую выполняет мышца в целом: Мышцы: Число мыш волокон: 1) Глаза 1) Менее 10 2)Пальцев рук 2) 1-25 3) Двуглавая 3) около 750 4) Камбаловидная 4) 2000 В мышцах, обеспечивающих наиболее точные и быстрые движения, двигательная единица состоит из нескольких мышечных воло кон, в то время как в мышцах, участвующих в поддержании позы, двигательные единицы включают несколько сотен и даже тысяч мышечных волокон. 25.14 Современная теория мышечного сокращения и расслабления. Теория мышечного сокращения: А. Электрохимическое преобразование: 1. Генерация ПД. 2. Распространение ПД по Т-системе.( по поперечной системе трубочек, которая служит связующим звеном между поверхностной мембраной и сократитель ным аппаратом мышечного волокна. ) 3. Электрическая стимуляция зоны контакта Т-системы и саркоплазматического ретикулума, активация ферментов, образование инозитолтрифосфата, повышение внутриклеточной концентрации ионов Са2+. Б. Хемомеханическое преобразование: 4. Взаимодействие ионов Са2+ с тропонином, освобождение ак тивных центров на актиновых филаментах. 5. Взаимодействие миозиновой головки с актином, вращение го ловки и развитие эластической тяги. 6. Скольжение нитей актина и миозина относительно друг друга, уменьшение размера саркомера, развитие напряжения или укоро чение мышечного волокна. Мышечное расслабление: Для расслабления в первую очередь необходимо по нижение концентрации ионов Са2+. Экспериментально было доказа но, что саркоплазматическая сеть имеет специальный механизм — кальциевый насос, который активно возвращает кальций в цистерны. Активация кальциевого насоса осуществляется неорганическим фос фатом, который образуется при гидролизе АТФ, а энергообеспечение работы кальциевого насоса также за счет энергии, образующейся при гидролизе АТФ. Таким образом, АТФ является вторым важнейшим фактором, абсолютно необходимым для процесса расслабления. Неко торое время после смерти мышцы остаются мягкими вследствие пре кращения тонического влияния мотонейронов (см. главу 4). Затем концентрация АТФ снижается ниже критического уровня и возмож ность разъединения головки миозина с актиновым филаментом исче зает. Возникает явление трупного окоченения с выраженной ригидно стью скелетных мышц. 26.16 Особенности строения и функционирования гладких мышц Гладкие мышцы находятся в стенке внутренних органов, крове носных и лимфатических сосудов, в коже н морфологически отли чаются от скелетной и сердечной мышц отсутствием видимой по перечной исчерченности.(тк миофибриллы располагаются хаотично) Гладкие мышцы состоят из клеток веретенообразной формы, сред няя длина которых 100 мкм, а диаметр 3 мкм. Клетки располагаются в составе мышечных пучков и тесно прилегают друг к другу. Мембраны прилежащих клеток образуют нексусы, которые обеспечивают электрическую связь между клетками и служат для передачи возбуж дения с клетки на клетку. Гладкие мышечные клетки содержат миофиламенты актина и миозина, которые располагаются здесь менее упорядоченно, чем в волокнах скелетной мышцы. Саркоплазматиче ская сеть в гладкой мышце менее развита, чем в скелетной. Особенности: 1) Статическая ( сохранительная, тоническая) 2) Низкая электропроводимость и высокая хемовозбудимость 3) Автоматия. ПД гладких мышечных клеток имеют авторитмиче ский (пейсмекерный) характер, подобно потенциалам проводящей системы сердца. 3) Низкая лабильность 4) Низкая проводимость 5) Высокая пластичность.Если растянуть висцеральную гладкую мышцу, то ее напряжение будет увеличиваться, однако если мышцу удерживать в состоянии удлинения, вызванным рас тяжением, то напряжение будет постепенно уменьшаться, иногда не только до уровня, существовавшего до растяжения, но и ниже этого уровня. 6) Слабые, практически неутомлямые 7) Реакция на растяжение. Уникальной особенностью висцеральной гладкой мышцы является ее реакция на растяжение. В ответ на растяжение гладкая мышца сокращается. 8) В механизме сокращения гладкой мышцы имеется особенность, отличающая его от механизма сокращения скелетной мышцы. Эта особенность заключается в том, что прежде чем миозин гладкой мышцы сможет проявлять свою АТФазную активность, он должен быть фосфорилирован. 9) Химическая чувствительность. Гладкие мышцы обладают высо кой чувствительностью к различным физиологически активным веще ствам: адреналину, норадреналину, АХ, гистамину и др. 27.17 Законы проведения возбуждения по нервам. Механизм проведения нервного импульса по безмиелиновым и миелиновым нервным волокнам. 1) Физиологическая целостность : для проведения возбуждения по нерву необходима не только его анатомическая целостность, но и физиологическая (физиол. Св-ва: возбу-ть, пров-ть, лабильность…) 2) Закон двустороннего проведения возбуждения: в нервном стволе возбуждение распространяется в обе стороны от места нанесения раздражения 3) Закон изолированного проведения возбуждения: возбуждение по нервным волокнам проводится изолированно 4) Закон относительной неутомляемости нерва(Введенский) вследствие того, что процессы ресинтеза энергии в нем идут с достаточно большой скоростью и успевают восстановить траты энергии, происходящие при прохождении возбуждения. В безмиелиновых волокнах возбуждение постепенно охватывает соседние участки мембраны осевого цилиндра и так распространяется до конца аксона. Скорость распространения возбуждения по волокну определяется его диаметром. В нервных безмиелиновых волокнах, где процессы метаболизма не обеспечивают быструю компенсацию расхода энергии на возбуждение, распространение этого возбуждения идет с постепенным ослаблением — с декрементом. Декрементное проведение возбуждения характерно для низкоорганизованной нервной системы. У высших животных благодаря прежде всего наличию миелиновой оболочки и совершенства метаболизма в нервном волокне возбуждение проходит, не затухая, бездекрементно. Этому способствуют наличие на всем протяжении мембраны волокна равного заряда и быстрое его восстановление после прохождения возбуждения. В миелиновых волокнах возбуждение охватывает только участки узловых перехватов, т. е. минует зоны, покрытые миелином. Такое проведение возбуждения по волокну называется сальтаторным (скачкообразным). В результате узловые перехваты являются наиболее возбудимыми и обеспечивают большую скорость проведения возбуждения. Время проведения возбуждения по миелиновому волокну обратно пропорционально длине между перехватами. Высокая прочность, надежность, экономичность. 28.17 Рецепторы органов чувств, понятие, классификация , основные свойства и особенности. Механизм возбуждения. Понятие функциональной мобильности. Рецептором называют генетически детерменированные макромолекулы, которые располагаются на субсинаптической мембране и взаимодействуют с молекулами медиатора как ключ с замком, запуская при этом рективную систему синапса. Рецепторы делят на четыре группы: механо-, термо-, хемо- и фоторецепторы. Каждую группу подраз деляют на более узкие диапазоны рецепции. Например, зрительные рецепторы делятся на воспринимающие освещенность, цвет, слухо вые — определенный тон, вкусовые — определенные вкусовые раздражения (соленое, сладкое, горькое) и т. д. Рецепторный потенциал возникает при раздражении рецептора как результат деполяризации и повышения проводимости участка его мембраны, который называется рецептивным. Рецептивный уча сток мембраны имеет специфические свойства, в том числе биохи мические, отличающие его от мембраны тела и аксона. Возникший в рецептивных участках мембраны рецепторный потенциал электротонически распространяется на аксонный холмик рецепторного нейрона, где возникает генераторный потенциал. Возникновение генераторного потенциала в области аксонного холмика объясняется тем, что этот участок нейрона имеет более низкие пороги возбуждения и потенциал действия в нем развивается раньше, чем в других частях мембраны нейрона. Чем выше генераторный потенциал, тем интенсивнее частота разрядов распространяющегося потенциала действия от аксона к другим отделам нервной системы. Следовательно, частота разрядов рецепторного нейрона зависит от амплитуды генераторного потенциала. Рецепторные нейроны различаются по скорости уменьшения их реакции (адаптации) на длящуюся стимуляцию. Рецепторные нейроны, медленно адаптирующиеся к раздражению, т. е. длительное время генерирующие потенциалы действия, называются тоническими. Рецепторы, быстро и коротко реагирующие на стимуляцию группой импульсов, называются физическими. Таким образом, реакция рецепторного нейрона, предназначенного для передачи информации из области восприятия, имеет 5 стадий: 1) преобразование сигнала внешнего раздражения; 2) генерация рецепторного потенциала; 3) распространение рецепторного потенциала по нейрону; 4) возникновение генераторного потенциала; 5) генерация нервного импульса. Мобильность функциональная — непрерывное изменение числа и возбудимости работающих рецепторов, обусловленное в основном влиянием на них со стороны ц.н.с. |