Лекция "Биологическое окисение"_2009. 2010г лекция 4 Тема Биологическое окисление этапы, цикл Кребса, пути использования кислорода в тканях
 Скачать 0.53 Mb. Скачать 0.53 Mb.
|

|
Факультеты: лечебно-профилактический, медико-профилактический, педиатрический. 2 курс. МИТОХОНДРИИ: ОСОБЕННОСТИ ХИМИЧЕСКОГО СОСТАВА, СТРОЕНИЯ.
Наружная мембрана содержит много белка порина, образующего гидрофильные каналы, которые пропускаю через мембрану неорганические ионы, метаболиты и даже небольшие белки (меньше 10кДа). Также она содержит ферменты: элонгазы (удлиняют молекулы насыщенных жирных кислот), кинуренингидроксилазу, моноаминооксидазу (маркер) и др. Межмембранное пространство митохондрий содержит аденилатциклазу, нуклеозиддифосфаткиназы. Внутренняя мембрана высокоспецифична, состоит на 70% из белков, которые выполняют каталитическую (окислительное фосфорилирование) и транспортную функцию. Внутренняя мембрана содержит ферменты: а). цепи окислительного фосфорилирования (цитохромоксидаза – маркер) б). СДГ в). β-оксибутират ДГ; г). карнитинацилтрансферазу, д). карнитинацилтранслоказу. Внутренняя мембрана на 30% состоит из фосфолипидов, из них 20% приходиться на кардиолипин, который делает мембрану непроницаемой для всех ионов. Матрикс на 50% состоит из белка и содержит сотни различных ферментов. Это ферменты: а). ЦТК; б). β-окисления жирных кислот; в). аминотрансферазы АСТ, АЛТ; г). глутамат ДГ д). фосфоенолпируваткарбоксилазу е). пируват ДГ. Также матрикс содержит несколько копий митохондриальной ДНК, митохондриальные рибосомы и тРНК. В клетке содержится от сотни до тысячи митохондрий, их размер 2-3 мкм в длину и 1 мкм в ширину. Метаболические и гомеостатические функции митохондрий В митохондриях происходит: синтез АТФ и теплопродукция в реакция окислительного фосфорилирования; β-окисления жирных кислот; реакции ЦТК, через ЦТК протекают некоторые реакции глюконеогенеза, переаминирования, дезаминирования, липогенеза и синтеза гема, осуществляется интеграция белкового, липидного и углеводного обмена. Причины и последствия повреждений митохондрий Повреждение внутренней мембраны митохондрий химическими и физическими факторами приводит разобщению окислительного фосфорилирования, нарушению синтеза АТФ, торможению анаболических реакций, межмембранного транспорта и всех видов обмена веществ. Оксидазный путь использования кислорода в клетке Оксидазный путь потребления кислорода протекает в митохондриях, потребляет 90% О2 и обеспечивает процесс окислительного фосфорилирования. Окислительным фосфорилированием называют синтез АТФ из АДФ и Н3РО4 за счет энергии движении электронов по дыхательной цепи. Окислительное фосфорилирование является основным источником АТФ в аэробных клетках. Хемиосмотическая теория Митчелла Для объяснения механизма окислительного фосфорилирования в 1961 году Митчеллом была предложена хемиосмотическая теория, которая включала четыре независимых постулата, касавшиеся функции митохондрий:
Современные представления о механизме окислительного фосфорилирования В настоящее время открыты все основные компоненты окислительного фосфорилирования, изучено их строение и свойства. Открыты основные принципы окислительного фосфорилирования, регуляция и механизмы некоторых стадий. МЕХАНИЗМ ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ Окислительное фосфорилирование состоит из процессов окисления и фосфорилирования, которые между собой сопряжены. Процесс окисления Процесс окисления происходит при движении электронов по дыхательной цепи от субстратов тканевого дыхания на кислород. Дыхательная цепь окислительного фосфорилирования состоит из 4 белковых комплексов, встроенных во внутреннюю мембрану митохондрий и небольших подвижных молекул убихинона и цитохрома С, которые циркулируют в липидном слое мембраны между белковыми комплексами. 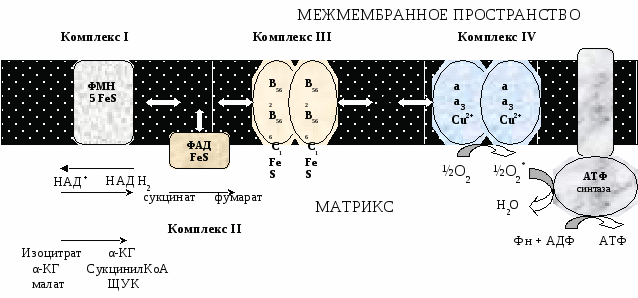 Комплекс I – НАДН2 дегидрогеназный комплекс – самый большой из дыхательных ферментных комплексов – имеет молекулярную массу свыше 800КДа, состоит из более 22 полипептидных цепей, в качестве коферментов содержит ФМН и 5 железо-серных (Fe2S2 и Fe4S4) белков. Комплекс II – СДГ. В качестве коферментов содержит ФАД и железо-серный белок. Комплекс III – Комплекс b-c1 (фермент QH2 ДГ), имеет молекулярную массу 500КДа, состоит из 8 полипептидных цепей, и вероятно существует в виде димера. Каждый мономер содержит 3 гема, связанных с цитохромами b562, b566, с1, и железо-серный белок. Комплекс IV – Цитохромоксидазный комплекс имеет молекулярную массу 300КДа, состоит из 8 полипептидных цепей, существует в виде димера. Каждый мономер содержит 2 цитохрома (а и а3) и 2 атома меди. Коэнзим Q (убихинон). Липид, радикал которого у млекопитающих образован 10 изопреноидными единицами (Q10). Убихинон переносит по 2Н+ и 2е-. 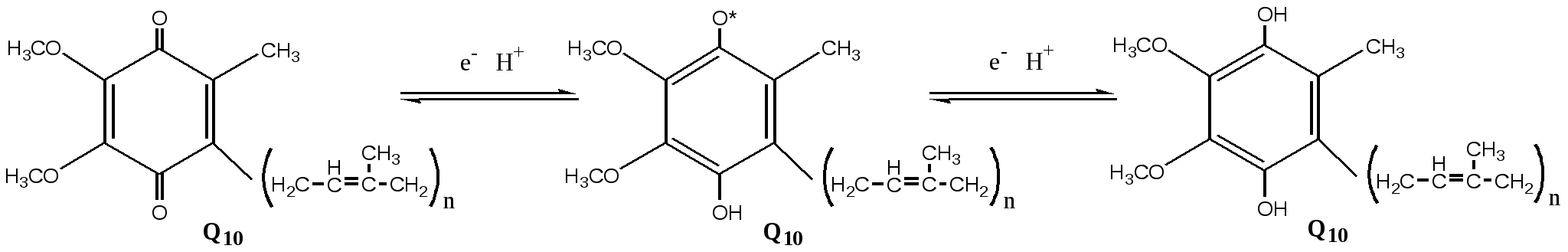 убихинон ↔ семихинон ↔ гидрохинон убихинон ↔ семихинон ↔ гидрохинонЦитохром с. Периферический водорастворимый мембранный белок с массой 12,5КДа, содержит 1 полипептидную цепь из 100 АК, и молекулу гема. Молекулярные соотношения между компонентами дыхательной цепи отличаются в разных тканях. Например, в миокарде, на 1 молекулу НАДН2 дегидрогеназного комплекса приходиться 3 молекулы комплекса b-c1, 7 молекул цитохромоксидазного комплекса, 9 молекул цитохрома С и 50 молекул убихинона. Электрохимический потенциал. Компоненты дыхательной цепи располагаются в мембране в порядке повышения их редокс-потенциала. При переходе е- от комплекса с низким редокс-потенциалом к комплексу с более высоким редокс-потенциалом происходит выделение свободной энергии. При окислении 1 НАДН2 выделяется 220 кДж/моль свободной энергии. I, III и IV комплексы дыхательной цепи используют 65-70% этой свободной энергии для переноса Н+ из матрикса митохондрий в межмембранное пространство, 30-35% свободной энергии рассеивается в виде тепла. 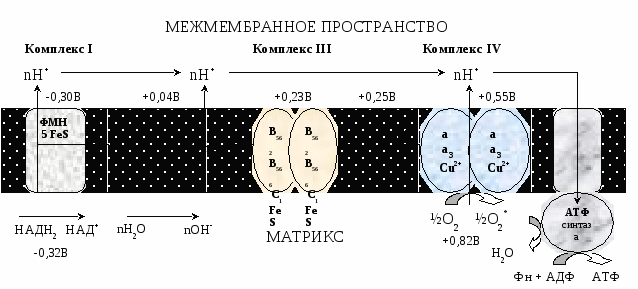 Этапы движения е- по дыхательной цепи Этапы движения е- по дыхательной цепи
При переносе Н+ из матрикса в межмембранное пространство на внутренней мембране создается осмотический градиент протонов ∆рН = 60 мВ (при ∆рН=1) (в матриксе рН выше, чем в цитозоле). Так как каждый Н+ несет положительный заряд, на внутренней мембране также появляется разность потенциалов ∆V=160мВ, внутренняя сторона мембраны заряжается отрицательно, внешняя – положительно. В сумме осмотический градиент протонов и разность потенциалов образуют электрохимический потенциал, который в типичной клетке составляет около 60+160=220 мВ. Механизм переноса Н+ через мембрану до конца не изучен. Вероятно, у разных компонентов дыхательной цепи существуют разные механизмы сопряжения транспорта е- с перемещением Н+. Образовавшийся на внутренней мембране митохондрий электрохимический потенциал используется для:
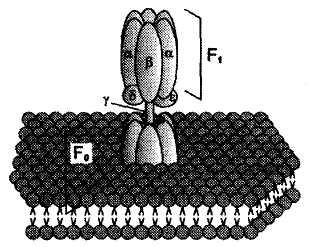
Процесс фосфорилирования Процесс фосфорилирования осуществляется АТФ-синтетазой (Н+-АТФ-аза), которая потребляет 40-45% свободной энергии, выделившейся при окислении. Н+-АТФ-аза интегральный белок внутренней мембраны митохондрий, она состоит из 2 белковых комплексов F0 и F1. 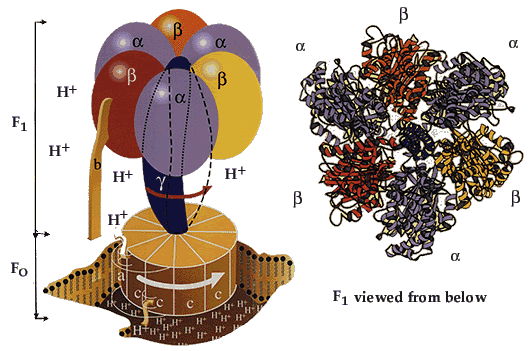
 АТФ-синтетаза обеспечивает обратимое взаимопревращение энергии электрохимического потенциала и энергии химических связей. Электрохимический потенциал внутренней мембраны заставляет Н+ двигаться из межмебранного пространства по каналу АТФ-синтазы в матрикс митохондрий. При каждом переносе протонов через канал Fo энергия электрохимического потенциала расходуется на поворот стержня, в результате которого циклически изменяется конформация а- и β-субъединиц и все 3 активных центра, образованных парам α- и β-субъединиц, катализируют очередную фазу цикла: 1) связывание АДФ и Н3РО4; 2) образование фосфоангидридной связи АТФ; 3) освобождение конечного продукта АТФ. Сопряжения и разобщения окислительного фосфорилирования Процесс окисления создает электрохимический потенциал, а процесс фосфорилирования его использует. Таким образом, электрохимический потенциал обеспечивает сопряжение (связывание) процессов окисления и фосфорилирования (окислительного фосфорилирования). Так как необходимый для сопряжения электрохимический потенциал создают I, III и IV комплексы дыхательной цепи, их называют пунктами сопряжения окисления и фосфорилирования. Повреждение внутренней мембраны митохондрий или увеличение ее проницаемости под действием разобщителей вызывает исчезновение электрохимического потенциала, разобщение процессов окисления и фосфорилирования, и прекращение синтеза АТФ. Разобщение дыхания и фосфорилирования называют явление исчезновения на мембране электрохимического потенциала под действием разобщителей и прекращение синтеза АТФ. Разобщителями являются вещества, которые могут переносить протоны (протонофоры) или другие ионы (ионофоры) через мембрану минуя каналы АТФ-синтетазы. В результате разобщения количество АТФ снижается, АДФ увеличивается, возрастает скорость потребления О2, окисления НАДН2, ФАДН2, а образовавшаяся свободная энергия выделяется в виде теплоты. Как правило, разобщители — липофильные вещества, легко проходящие через мембраны. Например, вещество 2,4-динитрофенол (переносит Н+), лекарство - дикумарол, метаболит - билирубин, гормон щитовидной железы - тироксин, антибиотики - валиномицин и грамицидин. Коэффициент окислительного фосфорилирования Для оценки эффективности окислительного фосфорилирования используют коэффициент окислительного фосфорилирования (Р/О). Коэффициентом окислительного фосфорилирования называют отношение количества фосфорной кислоты (Р), использованной на фосфорилирование АДФ, к атому кислорода (О), поглощённого в процессе дыхания. При окисление молекулы НАДН2, е- по дыхательной цепи проходят 3 пункта сопряжения, что обеспечивает синтез 3 АТФ при затрате 3 Н3РО4 и 3 АДФ на 1 атом кислорода. Соответственно для НАДН2 Р/О=3. При окисление молекулы ФАДН2, е- по дыхательной цепи проходят только 2 пункта сопряжения, что обеспечивает синтез 2 АТФ при затрате 2 Н3РО4 и 2 АДФ на 1 атом кислорода. Соответственно для ФАДН2 Р/О=2. Эти величины Р/О отражают теоретический максимум синтеза АТФ, фактически эта величина меньше из-за затрат на транспорт. Транспорт веществ через мембрану митохондрий 1). Внутренняя мембрана митохондрии проницаема для незаряженных простых молекул О2, Н2О, СО2, NH3, а также для монокарбоновых кислот. Эти вещества проходят мембрану самостоятельно по градиенту концентраций. 2). Для ионов мембрана не проницаема, через мембрану их переносят специальные насосы за счет энергии электрохимического потенциала. Только на транспорт АТФ и АДФ расходуется около четверти всей энергии электрохимического потенциала. Симпортом с Н+ в матрикс перемещается ПВК и Са2+. Антипортом перемещаются: 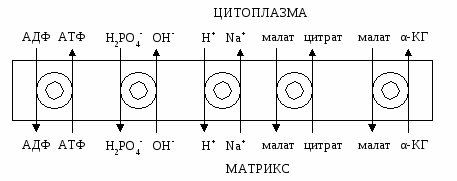 В результате обменных механизмов антипорта поддерживается осмотическое равновесие. Теплопродукция 30-35% свободной энергии рассеивается в виде теплоты и используется теплокровными животными на поддержание температуры тела. Кроме того, дополнительное образование теплоты может происходить при paзобщении дыхания и фосфорилирования в бурой жировой ткани. Она содержит много митохондрий с большим количеством дыхательных ферментов и разобщающего белка термогенина (РБ-1, около 10% всех белков). Разобщение окислительного фосфорилирования в бурой жировой ткани позволяет генерировать тепло для поддержания температуры тела у новорождённых, зимнеспящих животных и у всех млекопитающих в процессе адаптации к холоду. У взрослых людей важную роль в термогенезе при переохлаждении играют жирные кислоты. Охлаждение стимулирует выделение норадреналина, который активирует липазу в жировой ткани и гидролиз ТГ. Образующиеся свободные жирные кислоты способны не только быть топливом дыхательной цепи, но и переносить протоны через мембрану в матрикс митохондрий разобщая дыхание и фосфорилирование. Обратно жирные кислоты в ионизированной форме возвращаются с помощью переносчиков. Дыхательный контроль В норме субстраты тканевого дыхания и О2 находятся в достаточном количестве и не лимитируют окислительное фосфорилирование. Активность окислительного фосфорилирования ограничивает только концентрация АДФ, которая обратно пропорциональна концентрации АТФ. При нагрузке концентрация АТФ снижается, а АДФ увеличивается, что ускоряет дыхание и фосфорилирование. В состоянии покоя количество АТФ увеличивается, а АДФ снижается, что тормозит дыхание и фосфорилирование. Зависимость интенсивности дыхания митохондрий от концентрации АДФ называют дыхательным контролем. В результате дыхательного контроля скорость синтеза АТФ соответствует потребностям клетки в энергии. Общее содержание АТФ в организме 30—50 г, но каждая молекула АТФ в клетке «живёт» меньше минуты. В сутки у человека синтезируется 40—60 кг АТФ и столько же распадается. Макроэргические связи и макроэргические соединения, роль в организме. АТФ как важнейший аккумулятор и источник энергии В живых организмах существует целая группа органических фосфатов, гидролиз которых приводит к освобождению большого количества свободной энергии. Такие соединения называют высокоэнергетическими фосфатами. Разные фосфорилированные соединения обладают разным запасом свободной энергии. К группе высокоэнергетических фосфатов, помимо АТФ, относят енолфосфаты, ангидриды и фосфогуанидины. При гидролизе концевой фосфоангидридной связи АТФ превращается в АДФ и Рн. При этом изменение свободной энергии составляет —7,3 ккал/моль. Величина свободной энергии гидролиза АТФ делает возможным его образование из АДФ за счёт переноса фосфатного остатка от таких высокоэнергетических фосфатов, как, например, фосфоенолпируват или 1,3-бисфосфоглицерат; в свою очередь, АТФ может участвовать в таких эндергонических реакциях, как фосфорилирование глюкозы или глицерина. АТФ выступает в роли донора энергии в эндергонических реакциях многих анаболических процессов. Некоторые биосинтетические реакции в организме могут протекать при участии других нуклеозидтрифосфатов, аналогов АТФ; к ним относят гуанозинтрифосфат (ГТФ), уридинтрифосфат (УТФ) и цитидинтрифосфат (ЦТФ). Все эти нуклеотиды, в свою очередь, образуются при использовании свободной энергии концевой фосфатной группы АТФ. Наконец, за счёт свободной энергии АТФ совершаются различные виды работы, лежащие в основе жизнедеятельности организма, например, такие как мышечное сокращение или активный транспорт веществ. Таким образом, АТФ — главный, непосредственно используемый донор свободной энергии в биологических системах. В клетке молекула АТФ расходуется в течение одной минуты после её образования. У человека количество АТФ, равное массе тела, образуется и разрушается каждые 24 ч. Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счёт энергии окисления органических соединений. Цикл АТФ—АДФ — основной механизм обмена энергии в биологических системах, а АТФ — универсальная «энергетическая валюта». ЛЕКЦИЯ № 6 |