9.02 Погожий Гистология рыб. 37. Оплодотворение. Фазы оплодотворения. Контактные взаимодействия гамет. Активация спермия акросомная реакция. Активация яйцеклетки кортикальная реакция. Кариогамия
 Скачать 93.5 Kb. Скачать 93.5 Kb.
|

|
37. Оплодотворение. Фазы оплодотворения. Контактные взаимодействия гамет. Активация спермия - акросомная реакция. Активация яйцеклетки - кортикальная реакция. Кариогамия. Оплодотворение заключается в слиянии (сингамии) двух гамет разного пола — спермия (сперматозоида) и яйца. Оплодотворение имеет двоякое значение: 1) контакт спермия с яйцом выводит последнее из заторможенного состояния и побуждает к развитию; 2) слияние гаплоидных ядер спермия и яйца — кариогамия — приводит к возникновению диплоидного синкариона, объединяющего отцовские и материнские наследственные факторы. Возникновение при оплодотворении новых комбинаций этих факторов создаёт генетическое разнообразие, служащее материалом для естественного отбора и эволюции вида. Необходимая предпосылка оплодотворения — уменьшение числа хромосом вдвое, что происходит во время мейоза. Эти деления у мужских гамет осуществляются до формирования спермия, тогда как соотношения между делениями созревания яйца и оплодотворением у разных животных различно: спермий может проникать в яйцо до начала мейоза (губки, некоторые черви, моллюски, из млекопитающих — собака, лисица, лошадь); на стадии метафазы 1-го деления созревания (некоторые черви, моллюски, насекомые, асцидии); на стадии метафазы 2-го деления (ланцетник, многие позвоночные) и после завершения мейоза (кишечнополостные, морские ежи). Встреча сперматозоида с яйцом обычно обеспечивается плавательными движениями мужских гамет после того, как они выметаны в воду или введены в половые пути самки. Встрече гамет способствует выработка яйцами гамонов, усиливающих движения спермиев и продлевающих период их подвижности, а также веществ, вызывающих скопление спермиев вблизи яйца. Возникновение таких скоплений у гидроидных полипов из рода Campanularia и некоторых рыб иногда рассматривают как следствие привлечения спермиев (хемотаксис), подобного наблюдаемому при оплодотворении у мхов, папоротников и др. Однако существование направленных движений для спермиев животных не доказано; сперматозоиды двигаются беспорядочно и вступают в контакт с яйцом в результате случайного столкновения, а образование их скоплений, вероятно, вызывается действием механизма типа «ловушки», задерживающего спермии, случайно приблизившиеся к яйцу. Зрелое яйцо окружено оболочками, имеющими у некоторых животных отверстия для проникновения спермиев — микропиле. У большинства животных микропиле отсутствует, и, чтобы достигнуть поверхности ооплазмы, спермии должны проникнуть через оболочку, что осуществляется с помощью специального органоида сперматозоида — акросомы. После того как спермий концом головки коснётся яйцевой оболочки, происходит акросомная реакция: акросома раскрывается, выделяя содержимое акросомной гранулы, и заключённые в грануле ферменты растворяют яйцевые оболочки. В том месте где раскрылась акросома, её мембрана сливается с плазматической мембраной спермия; у основания акросомы акросомная мембрана выгибается и образует один или несколько выростов которые заполняются расположенным между акросомой и ядром (субакросомальным) материалом, удлиняются и превращаются в акросомные нити или трубочки. Длина этих нитей у разных животных варьирует от 1 до 90 мкм (в зависимости от толщины барьера, который спермию приходится преодолевать). Акросомная нить проходит через растворённую зону яйцевой оболочки, вступает в контакт с плазматической мембраной яйца и сливается с ней. У животных, спермии которых проникают в яйцо через микропиле (насекомые, головоногие моллюски, осетровые и костистые рыбы), акросома утрачивает своё первоначальное значение и иногда редуцируется или полностью исчезает (у некоторых веснянок, костистых рыб). У млекопитающих овулировавшее яйцо, кроме оболочки, окружено несколькими слоями фолликулярных клеток яйценосного бугорка. У лошади, коровы, овцы фолликулярные клетки рассеиваются вскоре после овуляции, и спермий свободно достигает поверхности яйцевой оболочки. У большинства млекопитающих клетки яйценосного бугорка сохраняются на протяжении нескольких часов и, чтобы проникнуть через этот барьер, спермии выделяют фермент гиалуронидазу, который растворяет вещество, связывающее фолликулярные клетки между собой. Гиалуронидаза, как и фермент, растворяющий яйцевую оболочку, заключена в акросоме. Сразу после эякуляции спермии неспособны к выделению этих ферментов; такая способность возникает под действием содержимого женских половых путей, вызывающего определённые физиологические изменения спермиев (процесс капацитации). С момента слияния плазматических мембран гамет в месте контакта акросомной нити с поверхностью ооплазмы яйцо и спермий — уже единая клетка — зигота. Вскоре обнаруживаются первые признаки активации яйца: кортикальная реакция и стягивание ооплазмы в месте контакта с акросомной нитью спермия, приводящее к образованию воспринимающего бугорка. Ооплазма этого бугорка обтекает ядро, центриоли и митохондрии сперматозоида, а иногда и осевой стержень его хвоста, вовлекая их в глубь яйца, тогда как плазматическая мембрана спермия остаётся на поверхности и встраивается в плазматическую мембрану яйца, так что поверхностная мембрана зиготы имеет мозаичное строение. К 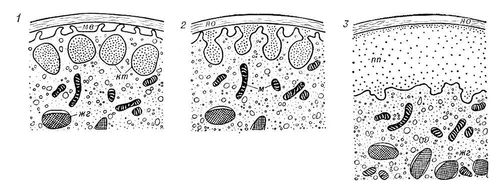 Рис. 1. Последовательные стадии кортикальной реакции (схема): 1 — зрелое яйцо до начала реакции; 2 — выделение содержимого кортекальных телец; 3 — содержимое кортикальных телец выделено и образовано приветеллиновое пространство; жг — желточные гранулы; кт — кортикальные тельца; м — митохондрии; мв — микроворсинки; пп — перивителлиновое пространство; яо — яйцевая оболочка. ортикальная реакция яйца (от лат. cortex — кора, скорлупа) ― это изменение кортикального (поверхностного) слоя яйца в ответ на активирующее воздействие. Распространяется волнообразно во все стороны от места контакта спермия с клеточной мембраной яйца или от места приложения искусственного активирующего воздействия (например, укола иглой). Видимым проявлениям кортикальной реакции предшествует латентный (скрытый) период, в течение которого по кортикальному слою яйца распространяется волна возбуждения (импульс активации яйца). Затем у большинства животных, в яйцах которых имеются кортикальные тельца, наступает видимая фаза кортикальной реакции: содержимое этих телец выделяется из яйца и оводняется, что приводит к отделению яйцевой оболочки от поверхности ооплазмы и образованию т. н. перивителлинового пространства (рис. 1). Кортикальная реакция охватывает всю поверхность яйца у морских ежей за 10—90 сек, у рыб за 2—5 мин (в зависимости от температуры). Кортикальная реакция играет важную роль в защите яйца от проникновения в него сверхчисленных спермиев (спермии агглютинируют при контакте с перивителлиновой жидкостью). Воздействия, тормозящие кортикальную реакцию, приводят к полиспермному оплодотворению. В результате кортикальной реакции и выделения из яйца веществ, локализованных в более глубоких слоях ооплазмы, изменяются свойства яйцевых оболочек и вокруг оплодотворённого яйца создаётся среда, благоприятная для его развития. Погрузившись в ооплазму, головка спермия поворачивается на 180, и у её основания формируется сперматическая звезда. Постепенно головка набухает и преобразуется в пузыревидный мужской пронуклеус, перемещающийся вслед за сперматической звездой, которая как бы увлекает его за собой. Мужской пронуклеус сближается с женским, а сперматическая звезда делится на две, участвующие затем в образовании веретена 1-го деления дробления. У кишечнополостных, плоских червей, морских ежей пронуклеусы сливаются в единое ядро зиготы, у некоторых круглых червей, моллюсков, ракообразных, рыб и земноводных они длительное время остаются в тесном контакте, но не сливаются, и объединение отцовского и материнского ядерного материала происходит только на стадии метафазы 1-го деления дробления. Одновременно с этими изменениями в яйце повышается интенсивность обмена веществ: увеличивается проницаемость клеточной мембраны, активируется синтез белка и др. Только этот вопрос страниц на 5 |