Микробиология, 1кт. микракт1. 5. Структурнофункциональная характеристика жгутиков, микроворсинок и включений. Методы их выявления
 Скачать 1.27 Mb. Скачать 1.27 Mb.
|

Особенности клеточной стенки грамположительных бактерийКлеточная стенка грамположительныхбактерийоднослойная, представ- лена 5–20рядамипептидогликана(составляет до 90 % сухой массы клеточной стенки). В пептидогликан погружены и выходят на поверхность тейхоевые кислоты (до 50 % ее сухой массы). Тейхоевые кислоты – растворимые в воде линейные полимеры, содержащие остатки глицерола или рибитола (глицерол- тейхоевые и рибитолтейхоевые кислоты). Тейхоевые кислоты – главные по-верхностные антигены многих грамположительных бактерий. Они в значи- тельном количестве располагаются междуцитоплазматическоймембраной и слоем пептидогликана, через поры в нем выступают наружу. Функция тейхо- евых кислот полностью не выяснена. Считают, что они придают муреиновому мешку определенную степень свободы при растяжении и сжатии, действуют наподобие пружины. Клеточная стенка большинства грамположительных бактерий не со-держит липидов, однако у микобактерий и коринебактерий в ней имеютсятоксические гликолипиды. Особенность пептидогликанов грамположитель-ных бактерий – частое отсутствие в них диаминопимелиновой кислоты.В клеточной стенке грамположительных бактерий отсутствуют липопо-лисахариды; содержание белка в них сильно варьирует. Белки во многом определяют антигенную специфичность таких бактерий. Например, стрепто- кокки серогруппы А по белкам М и Т подразделяют на несколько десятков серотипов. Особенности клеточной стенки грамотрицательных бактерий Клеточная стенка грамотрицательных бактерий значительнотоньше (14–18 нм) и состоит из двух слоев. Первый представлен одним или двумя рядами пептидогликана, на долю которого приходится до 5–10% сухой массы стенки. Для пептидогликана характерно низкое содержание попереч-ных сшивок между пептидными цепочками, однако в нем почти всегда при-сутствует мезо-диаминопимелиновая кислота. В клеточной стенке со-держитсямноголипопротеинов,фосфолипидов,липополисахарид,больше белка и, как правило, отсутствуют тейхоевые кислоты. Второйслой представлен бислоем фосфолипидов, пронизанным транспортными бел- ками, в нем крепится липополисахарид (ЛПС), включающий три компонента: липид А – крепит данную структуру в бислое фосфолипидов и проявляет токсичные свойства (эндотоксин); 2) ядро или остов – имеет одинаковое стро- ение у всех грамотрицательных бактерий; 3) О-специфическая цепь полисаха- рида – несет антигенную функцию (О-АГ). Клеточная стенка грамположительных бактерий плотно прилегает к ци- топлазматической мембране в отличие от клеточной стенки грамотрицатель- ных видов, компоненты которой (пептидогликановый слой и наружная мем- брана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп бактерий, харак- терно только для грамотрицательных форм. Периплазматическое пространство, или периплазма, содержит фер- менты (протеазы, липазы, фосфатазы, нуклеазы, бета-лактамазы), а также компоненты транспортных систем. Способность грамположительныхбактерийпри окраске по Граму удер- живать генциановый фиолетовый в комплексе с йодом (сине-фиолетоваяокраскабактерий) связана со свойством многослойного пептидогликана взаимодейство- вать с красителем. Кроме этого, последующая обработка мазка бактерий спиртом вызывает сужение пор в пептидогликане и тем самым задерживает краситель в клеточной стенке. Грамотрицательныебактериипосле воздействия спиртом утрачивают краситель, что обусловлено меньшим количеством пептидогликана (5–10 % массы клеточной стенки); они обесцвечиваютсяспиртоми при обра- ботке фуксином приобретаюткрасныйцвет. При нарушении синтеза клеточной стенки бактерий под влиянием лизо- цима, пенициллина, защитных факторов организма и других соединений обра- зуются клетки с измененной (часто шаровидной) формой: протопласты – бак- терии, полностью лишенные клеточной стенки, образуются обычно у грам- положительных бактерий; сферопласты – бактерии с частично сохранив- шейся клеточной стенкой, наблюдаются чаще у грамотрицательных бактерий, реже у грамположительных. Они не способны к размножению. После удале- ния ингибитора клеточной стенки такие измененные бактерии могут реверси- ровать, т.е. приобретать полноценную клеточную стенку и восстанавливать исходную форму. 7.Структурно-функциональная характеристика капсул и спор. Методы их обнаружения. Капсула, микрокапсула, слизь. Капсула – слизистая структура толщи- ной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имею- щая четко очерченные внешние границы. Состоит из полисахаридов (экзопо- лисахаридов), иногда из полипептидов; например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Она гидрофильна, вклю- чает большое (до 98 %) количество воды. Капсула различима в мазках-отпе- чатках из патологического материала. Поверхность колоний клеток с капсу- лами выглядит гладкой, влажной, блестящей. Многие бактерии образуют микрокапсулу – слизистое образование толщи- ной менее 0,2 мкм, выявляемое лишь при электронной микроскопии. От капсулы следует отличать слизь, состоящую из мукоидных экзополисахаридов и не имею- щую четких внешних границ. Слизь растворима в воде. Мукоидные экзополисахариды характерны для мукоидных штаммов си- негнойной палочки, часто встречающихся в мокроте больных с кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипа- нии к субстратам); их еще называют гликокаликсом. Кроме синтеза экзополи- сахаридов бактериями, существует и другой механизм их образования: путем действия внеклеточных ферментов бактерий на дисахариды. В результате этого образуются декстраны и леваны. Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют дей- ствию защитных факторов макроорганизма (фагоцитов, антител) и бакте- риофагов. Капсула антигенна(К-АГ): антитела против капсулы вызывают ее увеличение (реакция набухания капсулы). Капсула создает дополнительный осмотический барьер и является источником резервных питательных веществ. Споры – своеобразная форма покоящихся бактерий с грамположитель- ным типом строения клеточной стенки (рис. 12). Споры образуются при небла- гоприятных условиях существования бактерий (высушивание, дефицит питатель- ных веществ и др.). Внутри бактериальной клетки образуется одна спора (эндо- спора). Образование спор способствует сохранению вида и неявляетсяспособомразмножения,какугрибов. Форма спор может быть овальной или шаровидной. Расположение в клетке: терминальное – на конце палочки (у возбудителя столбняка); субтерминальное – ближе к концу палочки (у возбудителей ботулизма, газовой гангрены); центральное (у сибиреязвенной бациллы). 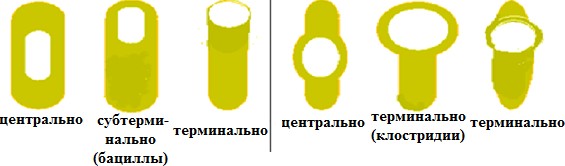 Спорообразование, форма и расположение спор в вегетативной клетке являются видовым свойством бактерий. Спорообразующие бактерии, у которых размер споры не превышает ши-рину клетки, называются бациллами (род Bacillus). Спорообразующие бакте- рии, у которых размер споры превышает ширину клетки, отчего они прини-маютформуверетена,называютсяклостридиями(род Clostridium, от лат. clostridium – веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля–Нильсена в красный, а вегетативная клетка – в синий цвет. Процесс спорообразования(споруляция) проходит ряд стадий, в течение ко- торых часть цитоплазмы и хромосома бактериальной вегетативной клетки отде- ляются, окружаясь врастающей цитоплазматической мембраной. Образуется проспора. Ее окружают две цитоплазматические мембраны, между которыми формируется толстый измененный пептидогликановый слой кортекса (коры). Изнутри он соприкасается с клеточной стенкой споры, а снаружи – с внутренней оболочкой споры. Наружная оболочка споры образована вегетативной клеткой (рис. 13, 14). 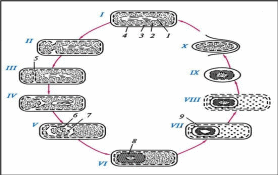 Рис. 13. Формирование эндоспоры спорообразующими бактериями: I – вегетативная клетка; II – инвагинация ЦПМ; III – образование споровой перего- родки (септы); IV – формирование двойной мембранной системы образующейся проспоры; V – сформированная проспора; VI – формирование кортекса; VII – формирование покровов споры; VIII – лизис материнской клетки; IX – свободная зрелая спора; X – прорастание споры; 1 – нуклеоид; 2 – цитоплазма; 3 – ЦПМ; 4 – клеточная стенка; 5 – споровая перего- родка; 6 – наружная мембрана споры; 7 – внутренняя мембрана споры; 8 – кортекс; 9 – покровы споры. 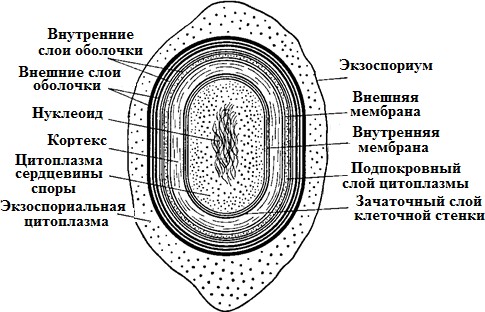 Споры некоторых бактерий имеют дополнительный покров – экзоспориум. Споры некоторых бактерий имеют дополнительный покров – экзоспориум.Таким образом формируется многослойная плохо проницаемая оболочка. Спорообразование сопровождается интенсивным потреблением проспорой, а за- тем формирующейся оболочкой споры дипиколиновой кислоты и ионов кальция. Спора приобретает термоустойчивость,которую связывают с наличием в ней дипиколината кальция. Спора может долго сохраняться во внешней среде из-за наличия много- слойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма. В почве, например, возбудители сибирской язвы и столбняка могут сохраняться десятки лет. Вблагоприятныхусловияхспорыпрорастают,проходятрипоследова-тельные стадии: 1) активацию; 2) инициацию; 3) прорастание. При этом из одной споры образуется одна бактерия. Активацияосуществляется при температуре 60–80°С. Также прорастание споры активируют глюкоза и другие углеводы, многие аминокислоты (в первую очередь L-аланин), а также некоторые ионы. Инициацияпрорастания необратима, она длится несколько минут. Под воз- действием аутолизинов происходят расщепление кортекса, поглощение воды и набухание (вздутие), в результате спора увеличивается. Стадия прорастанияхарактеризуется удалением из клетки дипиколиновой кислоты, ионов кальция, разрушением кортекса. В споре происходят глубокие физиологические изменения: усиливается дыхание, увеличивается активность ферментов. Активно синтезируются белок и РНК, реплицируется ДНК. Быстрый рост сопровождается разрушением оболочки и выходом проростка. В этот период спора утрачивает терморезистентность, снижается устойчивость споры к краскам, высушиванию. 8.Микроскопический метод исследования. Морфология бактерий. Изменение размеров микробных клеток. Микроскоп (от греч. μικρός– маленький и σκοπέω– смотрю) – оптический прибор для получения увеличенных изображений малых объектов, которые невозможно рассмотреть невооруженным глазом. До изобретения микроскопа для изучения мелких предметов использовали только лупу. В настоящее время достигнут предел разрешающей способности микро- скопа или микроскопии, равный длине опорной волны луча «жесткого» рент- геновского излучения, что соответствует длинам волн 1–10 нм. Для исследования объектов разного типа в зависимости от требуемой ве- личины оптического разрешения и других целей созданы разные микроскопы. |