9. Патология углеводного обмена лактазная недостаточность, галактоземия, фруктоземия, дефицит глюкозо6фосфатдегидрогеназы. Особенности диагностики и терапии у новорожденных, младенцев и детей раннего возраста
 Скачать 1.79 Mb. Скачать 1.79 Mb.
|

|
9.Патология углеводного обмена: лактазная недостаточность, галактоземия, фруктоземия, дефицит глюкозо-6-фосфатдегидрогеназы. Особенности диагностики и терапии у новорожденных, младенцев и детей раннего возраста Лактазная недостаточность (ЛН) – заболевание из группы ферментопатий. Развивается оно в том случае, если в организме фермент, отвечающий за расщепление молочного сахара (лактозы), либо полностью отсутствует, либо вырабатывается в недостаточном количестве. Диагностика:Важным в диагностике является молекулярно-генетическое исследование на предрасположенность к лактазной недостаточности. Анализ поможет в дифференциальной диагностике причин мальабсорбции лактозы и в подборе соответствующей диеты. ограничение продуктов, содержащих лактозу. Галактоземия (галактозный диабет) Причины галактоземии — генетически обусловленный дефект и/или недостаточность эффектов ферментов метаболизма галактозы. В связи с этим в плазме крови чрезмерно нарастает концентрация галактозы и ее метаболитов, оказывающих токсическое воздействие на нервную ткань, печень, почки, кишечник, клетки крови. Галактоземия связана с нарушением метаболизма галактозы вследствие недостаточности трех ферментов — галакто зо1- фос фа туридилтрасферазы (галактоземия I типа), галакто киназы (галактоземия II типа) и уридиндифосфат-галактозо-4-эпимиразы (галактоземия III типа). Все эти ферменты участвуют в превращении галактозы (входящей в состав «молочного сахара» лактозы) в глюкозу. При недостаточности указанных ферментов в организме в избытке накапливаются галактоза и ее метаболиты (галактозо-1-фосфат, галактотиол и др.). Они оказывают токсическое влияние на клетки головного мозга, печени, почек.Диагностика: У детей с галактоземией при рождении признаки заболевания отсутствуют. Симптоматика обнаруживается при кормлении ребенка грудным молоком или молочными смесями, содержащими лактозу. Первые проявления неспецифичны — срыгивания, задержка в прибавке массы тела, диарея. Фруктоземия, в т. ч. Врожденная непереносимость фруктозы Характеризуется недостаточностью фермента фрук то- 1-фосфатальдолазы и накоплением в клетках печени, почек, кишечника и других фруктозо-1-фосфата.Диагн;анализ крови,мочи. сключение фруктозы и сахарозы из рациона Дефицит глюкозо-6-фосфат-дегидрогеназы (Г6ФД) является наследственным генетическим заболеванием, которое может привести к разрушению эритроцитов (гемолиз). Г6ФД — это фермент, необходимый для поддержания стабильности гемоглобина. Гемоглобин — это белок, который содержится в эритроцитах и позволяет переносить кислород из легких и доставлять его во все части организма. Г6ФД помогает защитить гемоглобин от повреждений, вызванных стрессом от инфекций или использования определенных лекарственных препаратов или веществ. Люди с дефицитом Г6ФД наследуют дефект гена, отвечающего за выработку Г6ФД, поэтому в клетках крови у них недостаточно фермента. 8.Цикл пентозофосфатов. Значение в организме и взаимосвязь с другими видами обмена Пентозофосфа́тный путь— альтернативный путь окисления глюкозы, включает в себя окислительный и неокислительный этапы 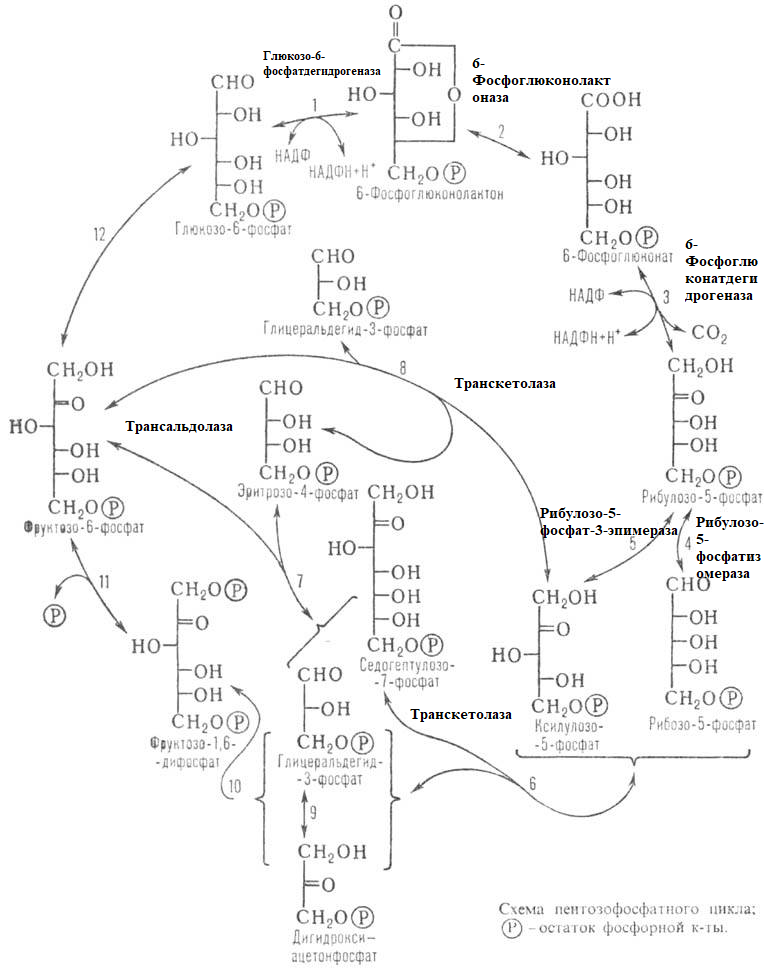 Суммарное уравнение окислительного этапа: Глюкозо-6-фосфат + 2 NADP+ + H2O → рибулозо-5-фосфат + 2 (NADPH + H+) + CO2. 6 глюкозо-6-фосфат + 12NADP + 2H2O → 12(NADPH + H+) + 5 глюкозо-6-фосфат + 6 СО2 Нарушения функционирования некоторых ферментов пентозофосфатного цикла приводят к развитию тяжелых заболеваний человека. Недостаточность глюкозо-6-фосфат-дегидрогеназы в эритроцитах служит причиной гемолитической анемии, а снижение активности транскетолазы в результате нарушения ее способности связывать тиамин приводит к развитию нервно-психического расстройства синдрома Вернике Корсакова 7.Особенности углеводного обмена в печени, мышечной и нервной тканях Углеводы – основные поставщики энергии в питании человека. Одной из важнейших функций печени в процессах обмена веществ является ее участие в поддержании постоянного уровня глюкозы в крови (глюкостатическая функция): глюкоза, поступающая в избытке, превращается в резервную форму, которая используется в период, когда пища поступает в ограниченном количестве. Энергетические потребности самой печени, как и других тканей организма, удовлетворяется за счет внутриклеточного катаболизма поступающей глюкозы. В печени катаболизм глюкозы представлен 2 процессами: 1) гликолитический путь превращения 1 моль глюкозы в 2 моль лактата с образованием 2 моль АТФ и 2) фосфоглюконатный путь превращения 1 моль глюкозы в 6 моль СО2 с образованием 12 моль НАДФ.Н. Оба процесса протекают в анаэробных условиях, обе ферментативные системы содержатся в растворимой части цитоплазмы, оба пути требуют предварительного фосфорилирования глюкозы. Гликолиз обеспечивает энергией клеточные реакции фосфорилирования, синтез белка; пентозофосфатный путь служит источником энергии восстановления для синтеза жирных кислот, стероидов. При аэробных условиях происходит сочетание гликолиза, протекающего в цитоплазме и цикла лимонной кислоты с окислительным фосфорилированием в митохондриях достигается максимальноый выход энергии в 38 АТФ на 1 моль глюкозы. Фосфотриозы, образующиеся в процессе гликолиза, могут быть использованы для синтеза α - глицерофосфата, необходимого для синтеза жиров. Пируват, который образуется при гликолизе, может быть использован для синтеза аланина, аспартата и других соединений, через стадию образования оксалоацетата. В печени реакции гликолиза могут протекать в обратном направлении и тогда происходит синтез глюкозы путем глюконеогенеза. В пентозофосфотном пути образуются пентозы, необходимые для синтеза НК. В отличие от гликолиза фосфоглюконатный путь необратим и здесь окисляется 1/3 глюкозы, 2/3 глюкозы окисляются по гликолитическому пути. В печени протекают гликогенез (биохимическая реакция, протекающая главным образом в печени и мышцах, во время которой глюкоза превращается в гликоген )и гликогенолиз( процесс расщепления гликогена до глюкозы).Эти процессы взаимосвязаны и регулируются как внутри – так и внеклеточными соотношениями между поступлением и потреблением глюкозы. 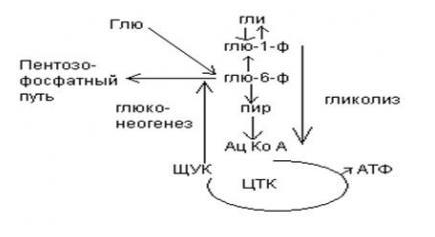 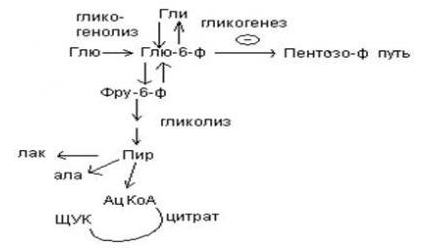 Печень учитывает запросы других органов и тканей в отношении углеводного обмена. В мышцах углеводный обмен происходит в соответствии с принципом самообслуживания. Цель мышечной клетки – наиболее эффективно использовать поступающую глюкозу для образования АТФ, необходимого для осуществления механической работы – сокращения. В состоянии покоя значительные количества глюкозы резервируются в форме гликогена. Цитоплазма мышечных клеток содержит в высоких концентрациях ферменты гликолиза, а изобилие митохондрий обеспечивает эффективный распад продуктов гликолиза через путь лимонной кислоты и цепь переноса электронов. Лишь в условиях крайнего утомления эти аэробные процессы е справляются с накоплением лактата. В мышцах идет гликогенез, мышца осуществляет лишь немногие синтетические функции. Ключевые ферменты глюконеогенеза в мышцах отсутствуют, и глюконеогенез не идет. Для востановительных синтезов в мышце НАДФ.Н не требуется, и фосфоглюконатный путь почти не функционирует. Обмен углеводов в мышцах обеспечивает создание тканевых запасов гликогена в состоянии покоя и использование этих запасов, а также поступающей глюкозы при напряженной работе; основные энергетические потребности всех типов мышц удовлетворяются главным образом за счет окисления продуктов обмена жиров. Ни медленно сокращающаяся гладкая мышечная ткань, ни сердечная мышца не потребляют глюкозу в значительной мере. Во время напряженной работы сердце обеспечивает себя лактатом для окисления. Фосфорилирование глюкозы в мышцах происходит под дейстием гексокиназы, в печени этот процесс катализируется глюкокиназой. Эти ферменты отличаются по Кm. Кm гексокиназы значительно ниже Кm глюкокиназы. Фермент мышц – гексокиназа участвует во внутриклеточной регуляции, т.е. этот фермент будет фосфорилировать глюкозу только до тех пор, пока глюкозо-6-ф используется в мышцах для гликолиза или образования гликогена. Другое важнейшее различие между тканью печени и мышцы состоит в отсутствии в мышцах фермента глюкозо-6-фасфатазы.Нервная ткань По сравнению со всеми органами тела функций мозга в наибольшей степени зависит от обмена углеводов. Если в крови, поступающей к мозгу, концентрация глюкозы становится вдвое ниже нормальной, то в течение нескольких секунд наступает потеря сознания, а через несколько минут – смерть. Для того чтобы обеспечить освобождение достаточного количества энергии, катаболизм глюкозы должен осуществляться в соответствии с аэробными механизмами; об этом свидетельствует даже более высокая чувствительность мозга к гипоксии, чем гипогликемии. Метаболизм глюкозы в мозге обеспечивает синтез нейромедиаторов, аминокислот, липидов, компонентов нуклеиновых кислот. Фосфоглюконатный путь функционирует в небольшой мере, обеспечивая НАДФ.Н для некоторых из этих синтезов. Основной катаболизм глюкозы в ткани мозга протекает по гликолитическому пути. Гексокиназа мозга имеет высокое сродство к глюкозе, что обеспечивает эффективное использование глюкозы мозгом. Активность ферментов гликолиза велика. Высокая активность митохондриальных ферментов цикла лимонной кислоты предотвращает накопление лактата в тканях мозга; большая часть пирувата окисляется до Ац-КоА. Небольшая часть Ац-КоА используется для образования нейромедиатора ацетилхолина. Основное количество Ац-КоА подвергается окислению в цикле лимонной кислоты и дает энергию. Метаболизм цикла Кребса используется для синтеза аспартата и глутамата. Эти аминокислоты обеспечивают обезвреживание аммиака в тканях мозга. Мозг содержит мало гликогена (0,1% от общего веса); этот запас расходуется очень быстро. 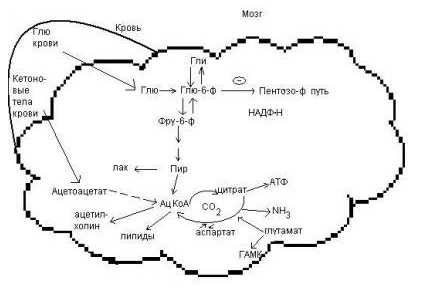 6.Глюконеогенез.Значение в организме и регуляция. Цикл Кори Глюконеогене́з — метаболический путь, приводящий к образованию глюкозы из неуглеводных соединений (в частности, пирувата). Наряду с гликогенолизом, этот путь поддерживает в крови уровень глюкозы, необходимый для работы многих тканей и органов, в первую очередь, нервной ткани и эритроцитов. Он служит важным источником глюкозы в условиях недостаточного количества гликогена, например, после длительного голодания или тяжёлой физической работы[1][2]. Глюконеогенез является обязательной частью цикла Кори, кроме того, этот процесс может быть использован для превращения пирувата, образованного при дезаминировании аминокислот аланина и серина Глюконеогенез протекает в основном в печени, но менее интенсивно он протекает также в корковом веществе почек[en] и слизистой кишечника. Глюконеогенез и гликолиз регулируются реципрокно: если клетка снабжена энергией в достаточной степени, то гликолиз приостанавливается, и запускается глюконеогенез; напротив, при активизации гликолиза происходит приостановление глюконеогенеза 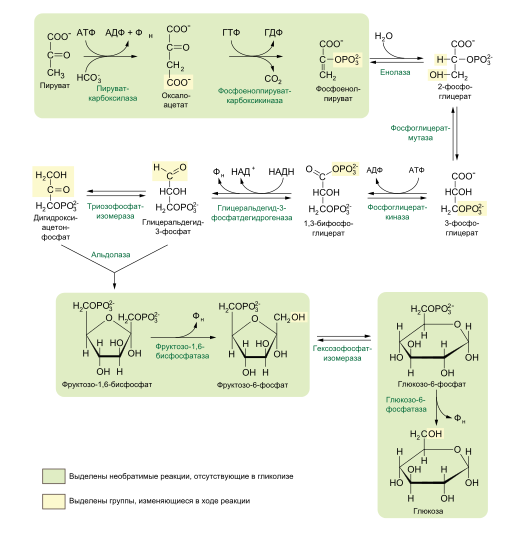 Ц 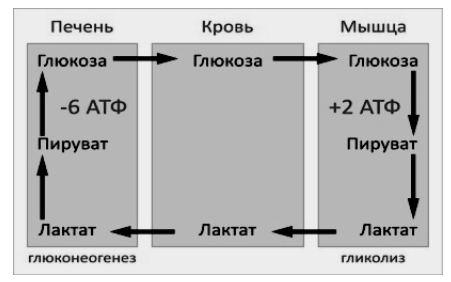 икл Кори (глюкозо-лактатный цикл). Он представляет собой биохимический транспорт лактата из мышц в печень и дальнейший синтез глюкозы из лактата, катализируемый ферментами глюконеогенеза икл Кори (глюкозо-лактатный цикл). Он представляет собой биохимический транспорт лактата из мышц в печень и дальнейший синтез глюкозы из лактата, катализируемый ферментами глюконеогенезаПри интенсивной мышечной работе и в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах) глюкоза подвергается анаэробному гликолизу с образованием лактата. При накоплении лактата в мышцах возникает лактат-ацидоз, раздражаются чувствительные нервные окончания, что вызывает боль в мышцах. Лактат переносится кровью в печень и превращается в пируват, а затем в глюкозу (глюконеогенез), которая с током крови Важность цикла основана на предотвращении лактоацидоза в мышцах в анаэробных условиях. Однако обычно, прежде чем это произойдет, молочная кислота перемещается из мышц в печень. Цикл также важен для производства АТФ, источника энергии, во время мышечной активности. может возвращаться в работающую мышцу. Направление лактатдегидрогеназной реакции в работающих мышцах и печени обусловлено отношением концентраций восстановленной и окисленной форм НАД+: отношение НАД+/НАДН в сокращающейся мышце больше, чем в печени. 5.Гликоген.Особенности структуры и биологическое значение. Реакция синтеза и распада. Регуляция . Гликогеновые болезни Гликоге́н — полисахарид состава (C6H10O5)n, образованный остатками глюкозы, соединёнными связями α-1→4 (в местах разветвления — α-1→6). В клетках животных служит основным запасным углеводом и основной формой хранения глюкозы. Откладывается в виде гранул в цитоплазме в клетках многих типов (главным образом в клетках печени и мышц Гликоген синтезируется в период пищеварения (через 1-2 ч после приёма углеводной пищи) Структура гликогена. А. Строение молекулы гликогена: 1 - остатки глюкозы, соединённые α-1,4-гликозидной связью; 2 - остатки глюкозы, соединённые α-1,6-гликозидной связью; 3 - нередуцирующие концевые мономеры; 4 - редуцирующий концевой мономер. Б. Строение отдельного фрагмента молекулы гликогена. Гликоген хранится в цитозоле клетки в форме гранул диаметром 10-40 нм. С гранулами связаны некоторые ферменты, участвующие в метаболизме гликогена, что облегчает их взаимодействие с субстратом. Разветвлённая структура гликогена обусловливает большое количество концевых мономеров, что способствует работе ферментов, отщепляющих или присоединяющих мономеры при распаде или синтезе гликогена, так как эти ферменты могут одновременно работать на нескольких ветвях молекулы. Гликоген депонируется главным образом в печени и скелетных мышцах. Синтез гликогена. 1 - глюкокиназа или гексокиназа; 2 - фосфоглюкомутаза; 3 - УДФ-глюкрпирофосфорилаза; 4 - гликогенсинтаза (глюкозилтрансфераза); 5 - фермент "ветвления" (амило-1,4 → 1,6-глюкозилтрансфераза), светлые и заштрихованные кружки - глюкозные остатки, закрашенные кружки - глюкозные остатки в точке ветвления. Распад гликогена или его мобилизация происходят в ответ на повышение потребности организма в глюкозе. Гликоген печени распадается в основном в интервалах между приёмами пищи, кроме того, этот процесс в печени и мышцах ускоряется во время физической работы. Распад гликогена происходит путём последовательного отщепления остатков глюкозы в виде глюкозо-1-фосфата. Гликозидная связь расщепляется с использованием неорганического фосфата, поэтому процесс называется фосфоролизом, а фермент гликогенфосфорилазой. Так же как и синтез, расщепление гликогена начинается с нередуцирующего конца полисахаридной цепи. При этом наличие разветвлённой структуры гликогена облегчает быстрое высвобождение глюкозных остатков, так как чем больше концов имеет молекула гликогена, тем больше молекул гликогенфосфорилазы могут действовать одновременно. Распад гликогена. В рамке - фрагмент гликогена с точкой ветвления. Закрашенный кружок - глюкозный остаток, связанный α-1,6-гликозидной связью; светлые и заштрихованные кружки - глюкозные остатки в линейных участках и боковых ветвях, связанные α-1,4-гликозидной связью. 1 - Гликогенфосфорилаза; 2 - олигосахаридтрансфераза; 3 - α-1,6-глюкозидаза. Обмен фенилаланина и тирозина. Особенности обмена тирозина в разных тканях. Синтез катехоламинов, меланинов, йодтиронинов. Наследственные биохимические блоки в распаде фенилаланина и тирозина: паркинсонизм, фенилкетонурия, алкаптонурия, альбинизм, диагностика и лечение. Фенилаланин - незаменимая аминокислота, так как в клетках животных не синтезируется её бензольное кольцо. Тирозин - условно заменимая аминокислота, поскольку образуется из фенилаланина. Содержание этих аминокислот в пищевых белках (в том числе и растительных) достаточно велико. Фенилаланин и тирозин используются для синтеза многих биологически 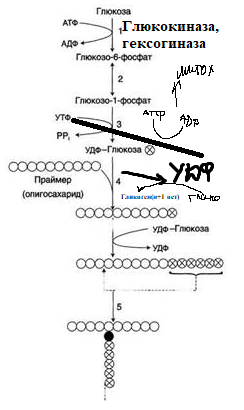 Гликогеновая болезнь, тип V (синонимы: болезнь Мак-Ардла, недостаточность мышечной фосфорилазы) - заболевание из группы наследственных болезней обмена веществ, обусловленное недостаточностью мышечной гликоген фосфорилазы, приводящей к накоплению гликогена в мышцах. Тип наследования - аутосомно-рецессивный. Гликогеновая болезнь, тип V (синонимы: болезнь Мак-Ардла, недостаточность мышечной фосфорилазы) - заболевание из группы наследственных болезней обмена веществ, обусловленное недостаточностью мышечной гликоген фосфорилазы, приводящей к накоплению гликогена в мышцах. Тип наследования - аутосомно-рецессивный.ГЛИКОГЕНОЗЫ Ранее гликогенозы классифицировались по номерам, однако в связи с открытием новых видов этих болезней появилось много разночтений. В настоящее время гликогенозы делят по патогенетическому признаку на печеночные, мышечные и смешанные формы. Следует отметить, что при гликогенозах количество гликогена не всегда изменено, изменения могут быть только в структуре его молекулы. Печеночные гликогенозы Самый частый гликогеноз I типа или болезнь фон Гирке обусловлен аутосомнорецессивным дефектом глюкозо-6-фосфатазы. Из-за того, что этот фермент есть только в печени и почках, преимущественно страдают эти органы, и болезнь носит еще одно название – гепаторенальный гликогеноз. Даже у новорожденных детей наблюдаются гепатомегалия и нефромегалия, обусловленные накоплением гликогена не только в цитоплазме, но и в ядрах клеток. Кроме этого, активируется синтез липидов с возникновением стеатоза печени. Так как фермент необходим для дефосфорилирования глюкозо-6-фосфата с последующим выходом глюкозы в кровь, у больных отмечается гипогликемия, и, как следствие, ацетонемия, метаболический ацидоз, ацетонурия. Гликогеноз III типа или болезнь Форбса-Кори или лимит-декстриноз – это ауто- сомно-рецессивный дефект амило-α-1,6-глюкозидазы, "деветвящего" фермента, гидролизующего α-1,6-гликозидную связь. Болезнь имеет более доброкачественное течение, и частота ее составляет примерно 25% от всех гликогенозов. Для больных характерна гепатомегалия, умеренная за4держка физического развития, в подростковом возрасте возможна небольшая миопатия. 4.Цикл трикарбоновых кислот. 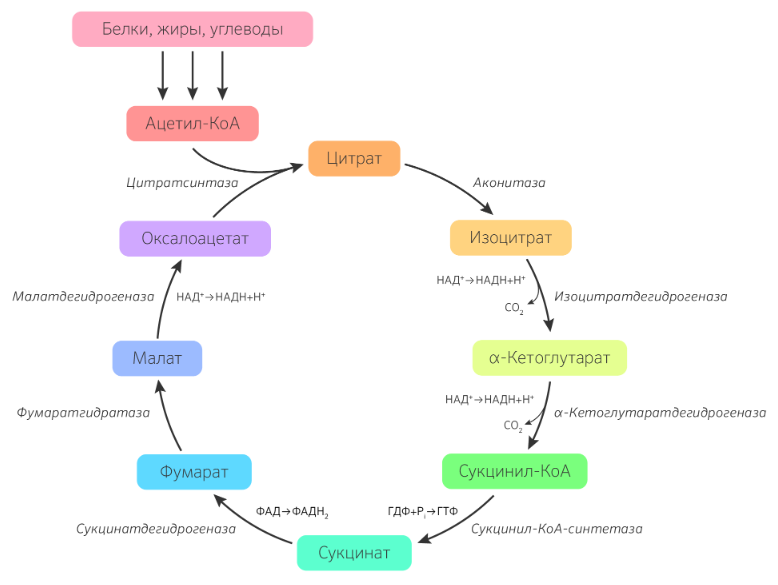 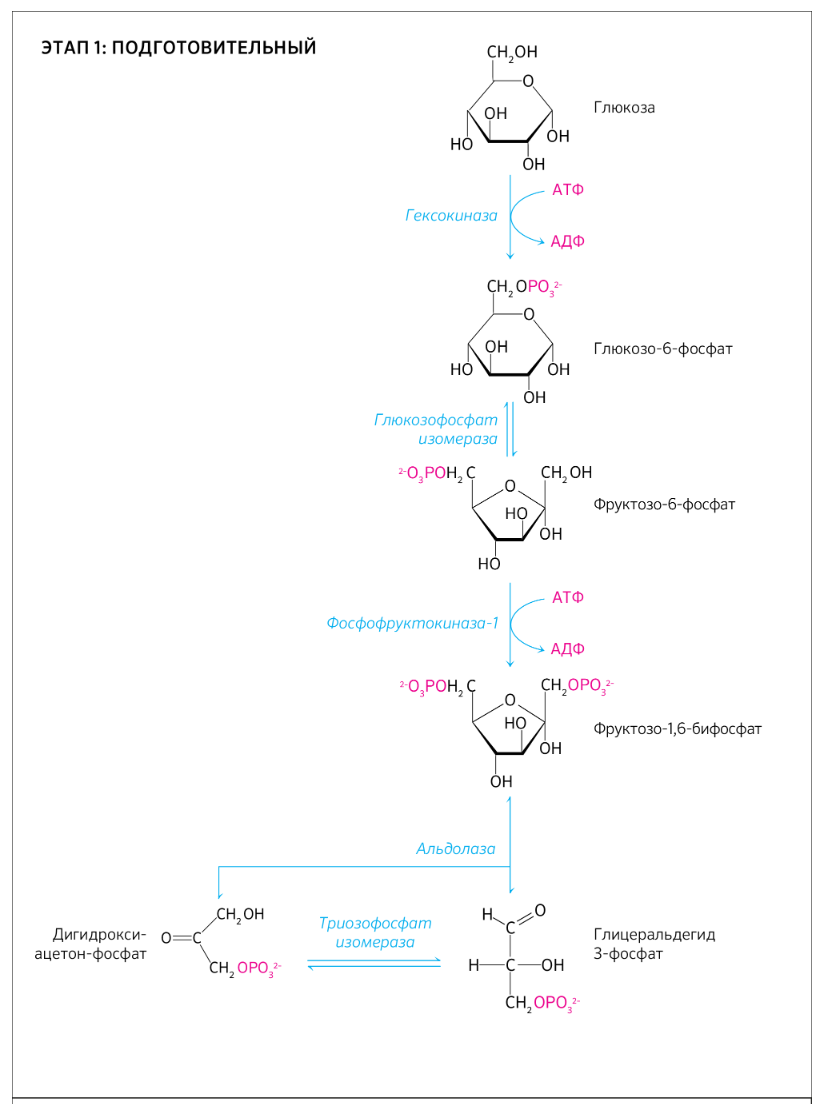 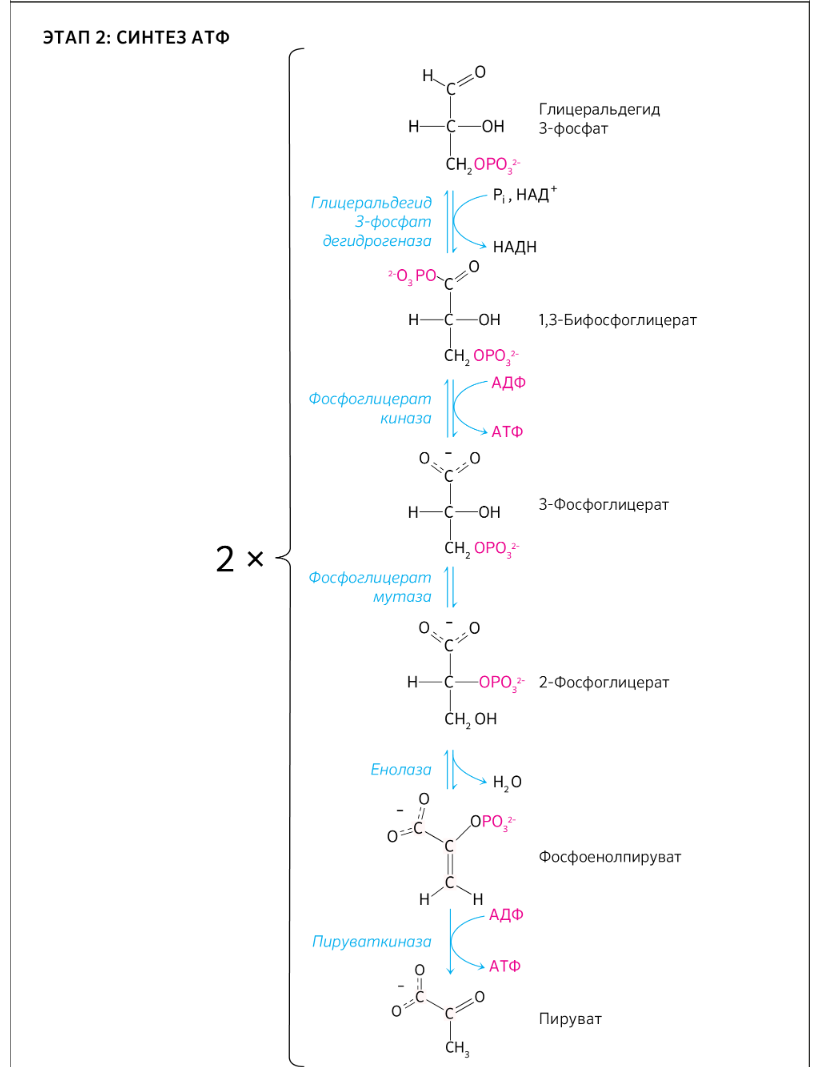 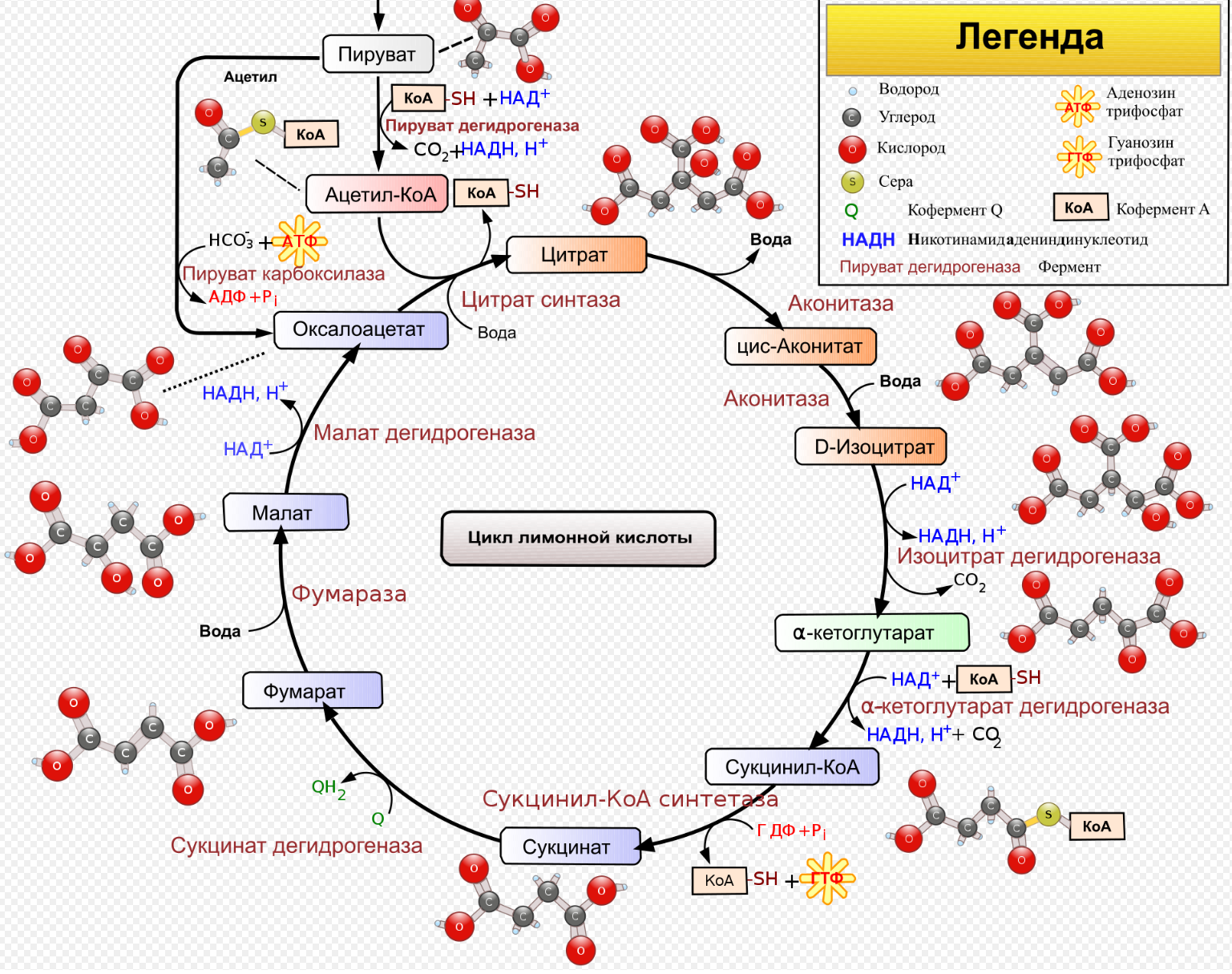 Цитосанитовая к-та  |