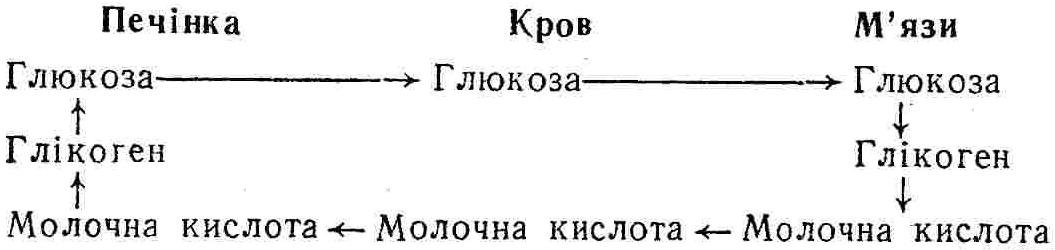лекция_8. АцетилКоА изолимонная кислота
 Скачать 1.9 Mb. Скачать 1.9 Mb.
|

50—70 %), потрапляє в клітини головного мозку, де окислюється до кінцевих продуктів. Усі енергетичні затрати центральної нервової системи, на відміну від інших органів, компенсуються за рахунок вуглеводів. Тому зниження вмісту цукру в крові негативно впливає насамперед на функції головного мозку. Інші органи і тканини організму також здатні поглинати глюкозу і здійснювати метаболічні перетворення її, однак провідна роль у цих процесах належить печінці.Перетравлювання і всмоктування вуглеводів. Глікогенна функція печінки. Анаеробне перетворення вуглеводів: глікогеноліз; гліколіз; спиртове бродіння. Аеробне перетворення вуглеводів: окислювальне декарбоксилювання піровиноградної кислоти; цикл трикарбонових кислот (Кребса). Биохимия / Под ред. В. В. Меньшикова, Н. И. Волкова. – М. : Физкультура и спорт, 1986. – 384 с. Проскурина И. К. Биохимия : учеб. пособие / И. К. Проскурина. – М. : ВЛАДОС-ПРЕСС, 2003. – 240 с. Роман С. В. Біохімія : лабораторний журнал (для студентів нехімічних (небіологічних) спеціальностей педагогічного університету) / С. В. Роман, Л. М. Крючок. – Луганськ : Альма-матер, 2008. – 36 с. Химия и биологическая химия : учеб. пособие / Явоненко А. Ф., Яковенко Б. В., Крутовский С. В. [и др.]. – К. : Вища школа, 1988. – 415 с. Яковлев Н. Н. Биохимия / Н. Н. Яковлев. – М. : Физкультура и спорт, 1974. – 320 с. Боєчко Ф. Ф. Біологічна хімія / Ф. Ф. Боєчко. – К. : Вища школа, 1995. – 536 с. Збарский Б. И. Биологическая химия / Збарский Б. И., Иванов И. И., Мардашев С. Р. – Л. : Медицина. Ленинградское отделение, 1972. – 582 с. Кильчевская М. А. Метаболический атлас / М. А. Кильчевская. – Минск : «Вышейшая школа», 1976. – 200 с. Мусил Я. Современная биохимия в схемах : пер. с англ. / Мусил Я., Новакова О., Кунц К. – М. : Мир, 1984. – 216 с. Савицкий И. В. Биологическая химия / И. В. Савицкий. – К. : Вища школа. Головное изд-во, 1982. – 472 с. Филиппович Ю. Б. Основы биохимии / Ю. Б. Филиппович. – М. : Изд-во «Агар», 1999. – 503 с. Біосинтез полісахариду глікогену (глікогенез). Процес синтезу глікогену інтенсивно проходить у печінці і має важливе значення в утворенні рухомого резерву полісахаридів в організмі. Вихідними спорками для синтезу глікогену можуть бути глюкоза, яка всмоктується з кишок у кров, а також глюкоза і глюкозо-6-фосфат, що утворюються в процесі глюконеогенезу. Процес синтезу глікогену розпочинається з фосфорилювання глюкози за участю ферменту гексокінази. При цьому утворюється глюкозо-6-фосфат: 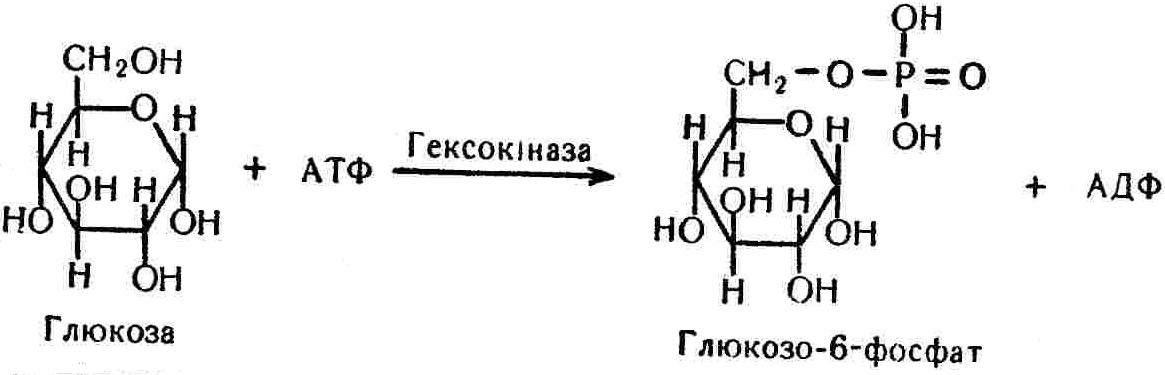 Останній під впливом фосфоглюкомутази перетворюється на глюкозо-1-фосфат:  Далі внаслідок взаємодії глюкозо-1-фосфату з уридинтрифосфорною кислотою утворюється уридиндифосфоглюкоза. 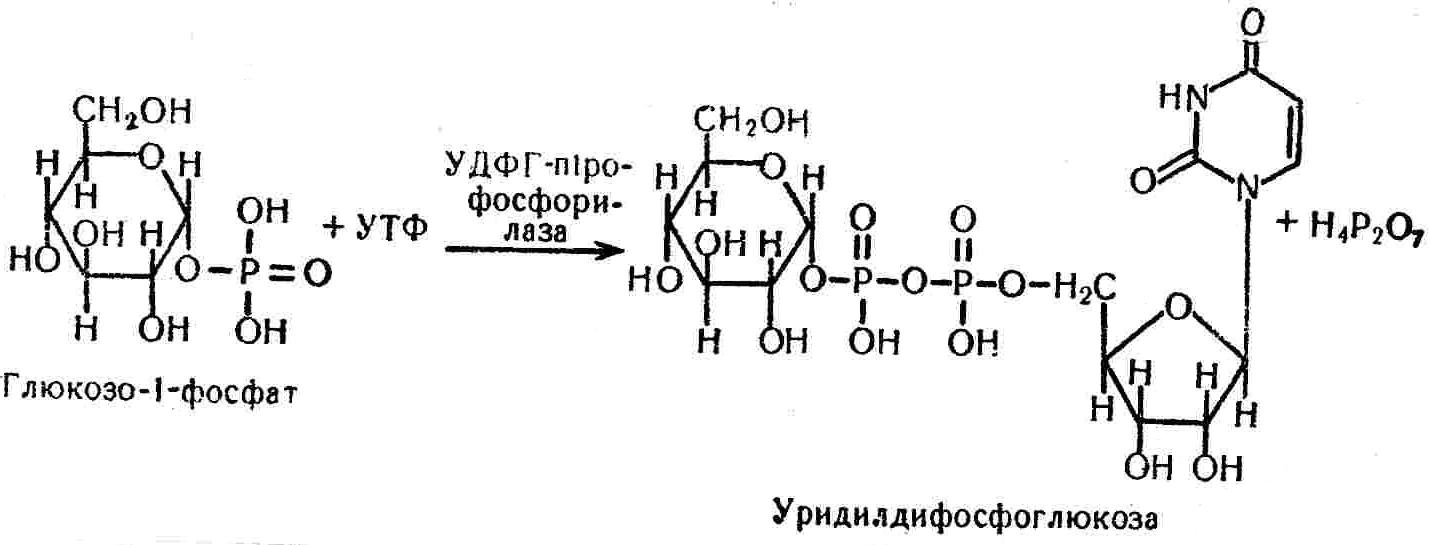 Реакція проходить під каталітичною дією ферменту глюкозо-1-фосфатуридилтрасферази (УДФГ-пірофосфорилази). На наступній стадії залишок глюкози від УДФ переноситься на ланцюг глікогену (затравку), внаслідок чого утворюється α(14)-глюкозидний зв'язок між першим атомом вуглецю залишку глюкози ланцюга глікогену і четвертим гідроксилом залишку глюкози. Реакцію каталізує глікогенсинтетаза: 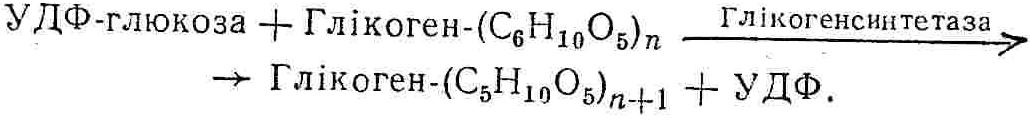 УДФ, що утворився внаслідок цієї реакції, піддається фосфорилюванню за участю АТФ і включається в новий цикл перетворень глюкозо-1-фосфату. Утворення α (16)-зв'язків у молекулі глікогену відбувається за участю спеціального розгалужуючого ферменту, внаслідок чого виникає розгалужена молекула глікогену, тобто глікоген, у якого поряд з α (14)-зв'язками в окремих місцях ланцюга є α(16)-зв'язки. Розкладання вуглеводів в організмі. Вуглеводи органів і тканин організмів людини і тварин, здебільшого глікоген і глюкоза, постійно зазнають різних перетворень, серед яких особливе місце належить процесам розкладання вуглеводів з вивільненням енергії. Розкладання вуглеводів у тканинах організмів людини і тварин відбувається анаеробно (за відсутності кисню) і аеробно (при наявності кисню). У першому випадку кінцевим продуктом є молочна кислота: 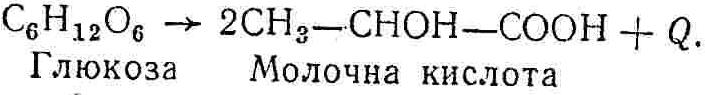 Під час аеробного розкладання вуглеводів кінцевими продуктами є СO2 і Н2О: 3. АНАЕРОБНЕ ПЕРЕТВОРЕННЯ ВУГЛЕВОДІВ Анаеробне перетворення вуглеводів проходить у різних органах і тканинах вищих організмів, а також у клітинах бактерій та мікроорганізмів. Основним субстратом для цього процесу є глюкоза і глікоген, а для окремих бактерій — пентози, жирні кислоти та амінокислоти. Анаеробне перетворення вуглеводів в організмах людини і тварин може розпочинатися з глікогену або глюкози. У першому випадку його називають глікогенолізом, у другому — гліколізом. У скелетних м'язах ці два процеси виражені в однаковій мірі, а в головному мозку і серцевому м'язі переважає гліколіз. Обидва процеси розпочинаються з перетворення глюкозо-6-фосфату, проходять однаково і каталізуються одними і тими самими ферментами, локалізованими у розчинній частині цитоплазми. Відмінність між цими процесами полягає лише у способі утворення фосфорильованої форми гексози — глюкозо-6-фосфату. Глікогеноліз. Процес розпочинається з фосфоролізу глікогену за участю ферменту глікогенфосфорилази. За цих умов внаслідок трансферазної реакції — перенесення глікозильного залишку з глікогену на фосфорну кислоту — утворюється глюкозо-1-фосфат (ефір Корі): 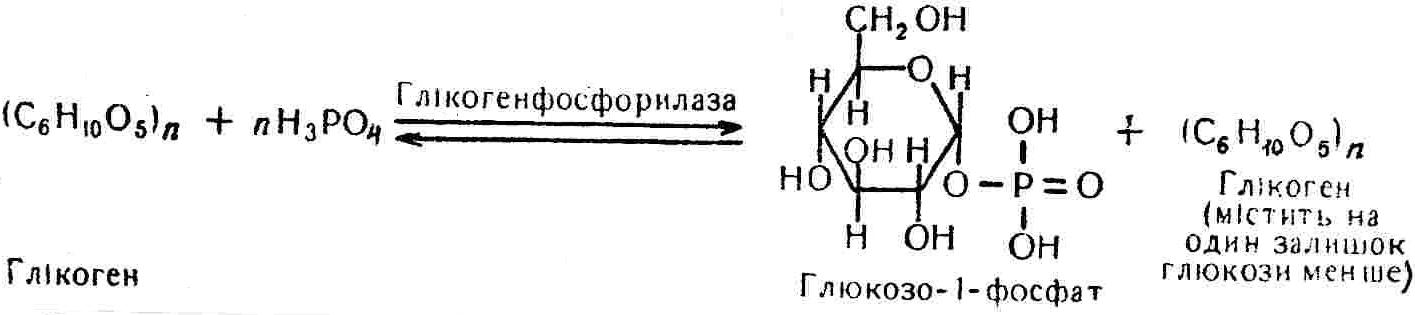 Глюкозо-1-фосфат, що утворився під впливом фосфорилази, перетворюється на глюкозо-6-фосфат. Реакцію каталізує фосфоглюкомутаза:  Далі перетворення глюкозо-6-фосфату відбувається так само, як і в процесі гліколізу. Гліколіз. Процес розпочинається з фосфорилювання глюкози за участю АТФ і ферменту гексокінази. При цьому утворюється також глюкозо-6-фосфат: Фосфорилювання глюкози є досить важливим етапом, оскільки лише в фосфорильованому стані вона здатна вступати в наступні метаболічні перетворення. Враховуючи це, реакція фосфорилювання глюкози дістала назву «шлюзової», або реакції «запалу». Вивільнення значної кількості енергії в ході гексокіназної реакції сприяє зміщенню рівноваги вправо і робить реакцію практично незворотною. Наступною реакцією гліколізу є ізомеризація глюкозо-6-фосфату в фруктозо-6-фосфат. Реакцію каталізує фермент глюкозо-6-фосфат-ізомераза:  Реакція протікає в основному в зворотному напрямку. У тканинах організму між двома фосфорними ефірами гексоз встановлюється динамічна рівновага (70 % глюкозо-6-фосфату і 30% фруктозо-6-фосфату). На наступному етапі фруктозо-6-фосфат за участю АТФ, іонів Mg2+ та ферменту фосфофруктокінази перетворюється на фруктозо-1,6-дифосфат (ефір Іванова): 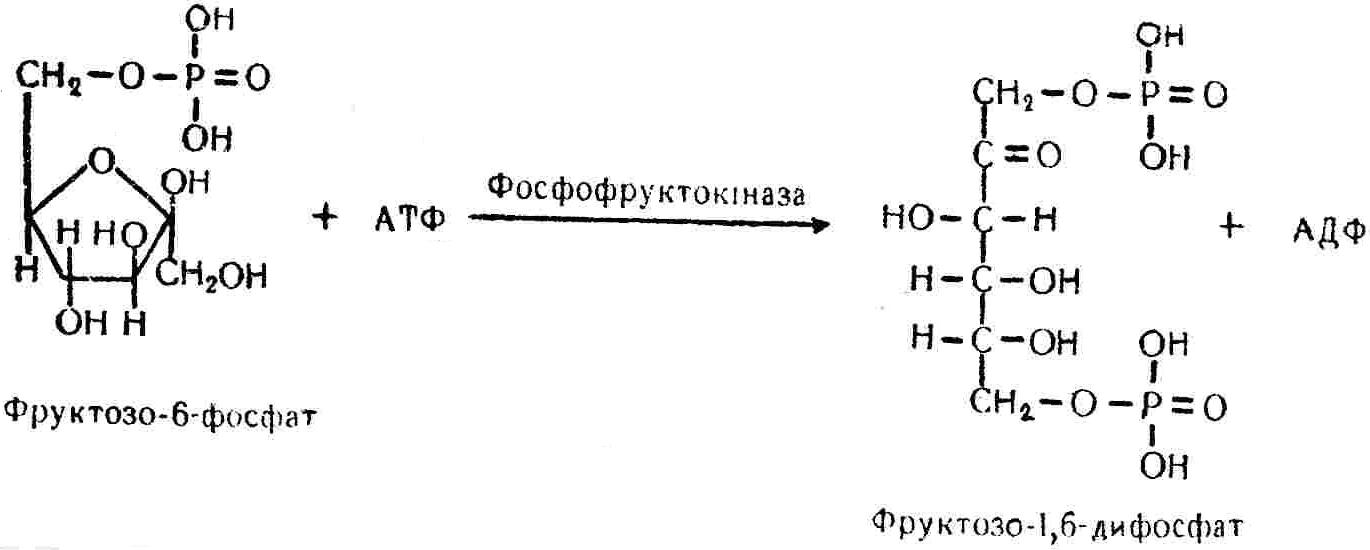 Реакція необоротна і є найповільнїшою у процесі гліколізу. Фермент фосфофруктокіназа активується АДФ і АМФ та інактивується при значному підвищенні вмісту АТФ. Отже, при інтенсифікації процесів окислювального фосфорилювання, коли збільшується відношення АТФ/АДФ, активність фосфофруктокінази послаблюється, що супроводжується сповільненням гліколізу. І, навпаки, при зменшенні відношення АТФ/АДФ процес гліколізу прискорюється. Наступний етап гліколізу забезпечує розкладання фруктозо-1,6-дифосфату за участю ферменту альдолази на дві молекули фосфотріоз – 3-фосфогліцериновий альдегід і діоксіацетонфосфат: 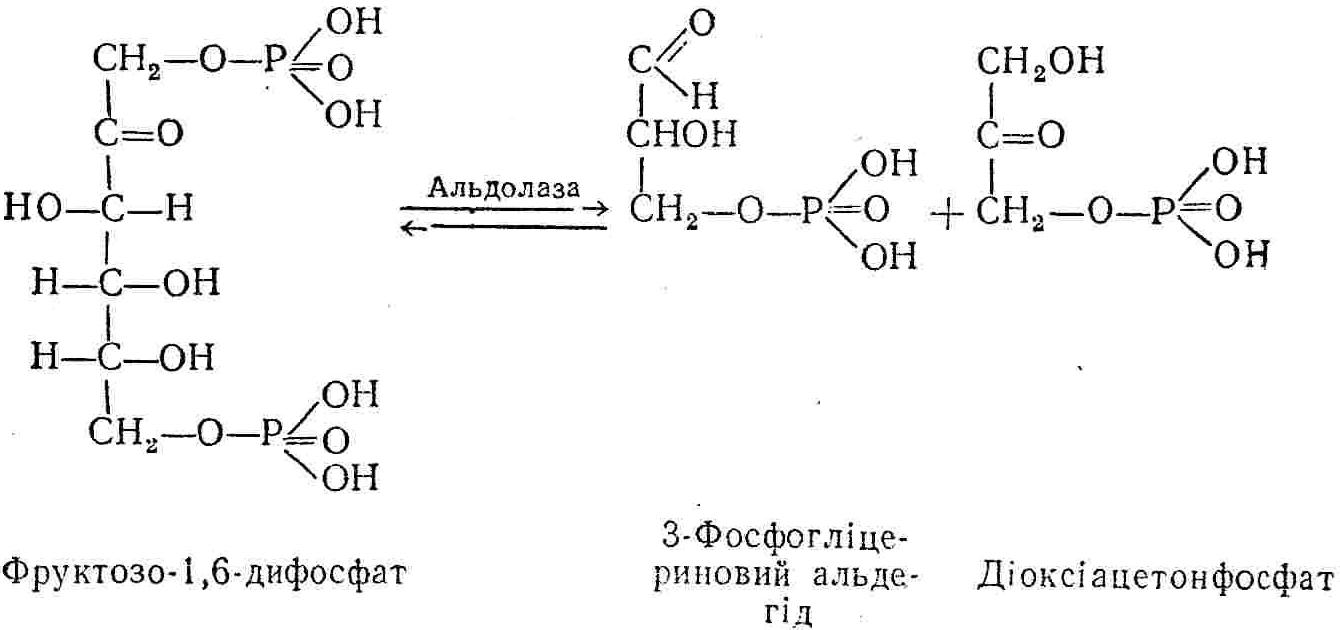 Далі частина діоксіацетонфосфату, що утворився, за участю ферменту тріозофосфатізомерази перетворюється на 3-фосфогліцериновий альдегід:  Отже, з кожної молекули глюкози утворюються дві молекули 3-фосфогліцеринового альдегіду. На цьому завершується перша стадія гліколізу, для якої характерне використання енергії АТФ. Її називають підготовчою стадією. Далі розпочинається друга стадія гліколізу, що супроводжується вивільненням енергії та акумулюванням її в макроергічних зв'язках АТФ. На наступній стадії гліколізу 3-фосфогліцериновий альдегід починає перетворюватись на 3-фосфогліцеринову кислоту. Реакція проходить у кілька етапів за участю специфічної дегідрогенази та неорганічного фосфату. 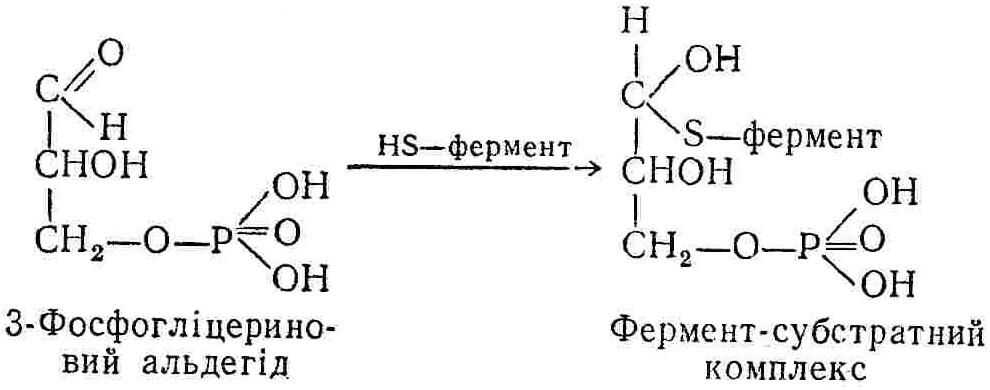 Молекула ферменту 3-фосфогліцеринальдегіддегідрогеназа складається з 4 субодиниць, кожна з яких містить 1 молекулу НАД+ і 4 вільних тіолових групи (НS-групи) залишків цистеїну, які входять до складу активного центра ферменту. В процесі каталітичної реакції на першому етапі 3-фосфогліцериновий альдегід зв'язується з молекулою ферменту за участю його тіолової групи з утворенням фермент-субстратного комплексу. На наступному етапі при наявності неорганічного фосфату 3-фосфогліцеринальдегіддегідрогеназа за участю НАД+ окислює фермент-субстратний комплекс. Енергія, що виділяється внаслідок реакції окислення, зосереджується в карбоніл-тіоловому зв'язку: 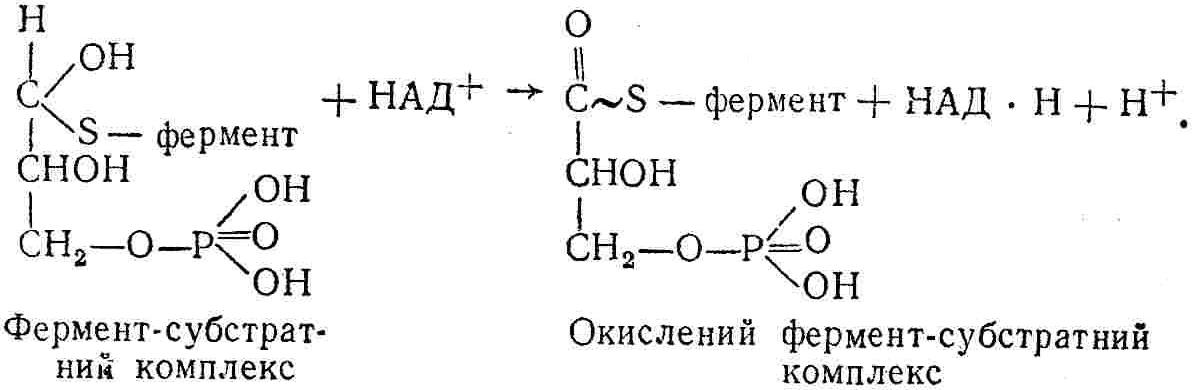 Окислений фермент-субстратний комплекс нестійкий, він взаємодіє з фосфорною кислотою, внаслідок чого утворюється 1,3-дифосфогліцеринова кислота. При цьому енергія карбоніл-тіолового зв'язку зосереджується на карбоксил-фосфатному зв'язку: 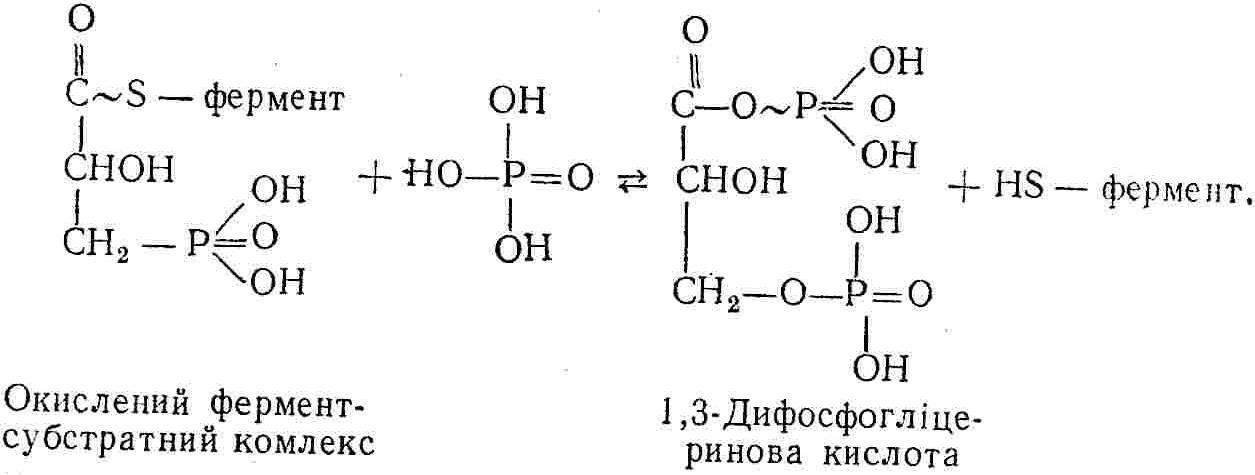 Наступна стадія перетворення 1,3-дифосфогліцеринової кислоти — перенесення від неї залишку фосфорної кислоти і енергії, зосередженої на карбоксил-фосфатному зв'язку, на АДФ. Внаслідок реакції утворюється 3-фосфогліцеринова кислота і АТФ. Реакцію каталізує фермент фосфогліцераткіназа: 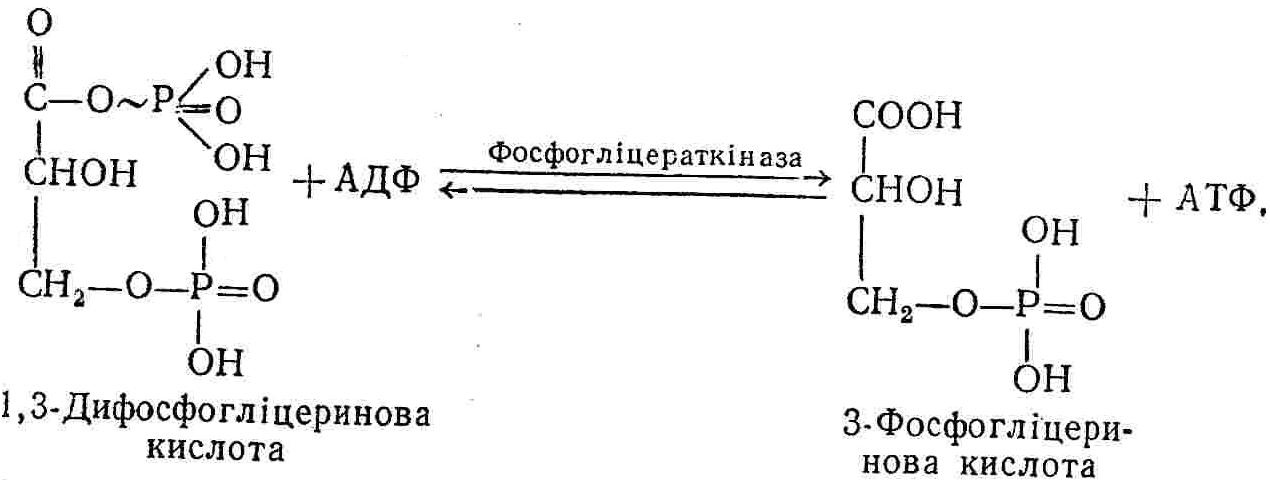 3-Фосфогліцеринова кислота, що утворилась, внаслідок каталітичної дії ферменту фосфогліцеромутази перетворюється на 2-фосфогліцеринову кислоту: 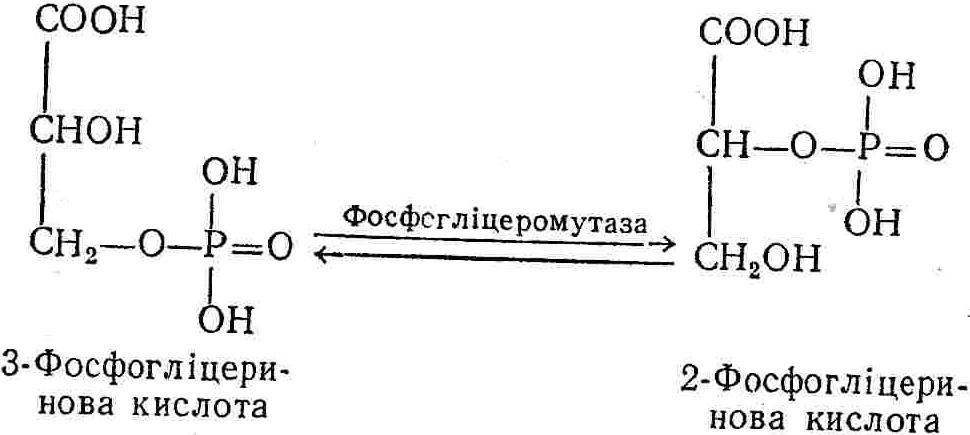 Остання, втрачаючи молекулу води, під впливом ферменту фосфопіруватгідратази перетворюється на 2-фосфоенолпіровиноградну кислоту. Ця реакція важлива в зв'язку з енергетичними змінами, які відбуваються під час її перебігу. Залишок фосфорної кислоти, що входить до складу 2-фосфогліцеринової кислоти, сполучений з останньою простим енергетично бідним ефірним зв'язком. Внаслідок енолізації відбувається перерозподіл внутрішньомолекулярної енергії і цей зв'язок перетворюється на макроергічний:  Далі 2-фосфоенолпіровиноградна кислота дефосфорилюється за участю АДФ, внаслідок чого енергетично багатий фосфатний залишок переходить до АДФ. Продуктом реакції є енольна форма піровиноградної кислоти, яка спонтанно перетворюється на піровиноградну кислоту і АТФ: 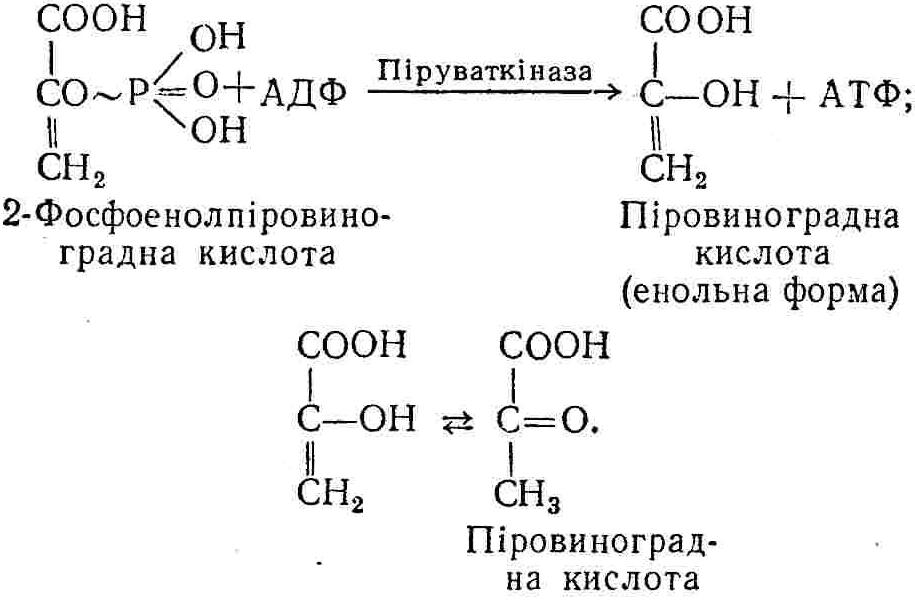 Реакцію каталізує фермент піруваткіназа, яка активується іонами Мg2+, Мn2+ або іонами лужних металів. Далі піровиноградна кислота, що утворилась, за участю ферменту лактатдегідрогенази перетворюється на молочну кислоту. Донором водню у цій реакції є відновлена форма коферменту НАД: 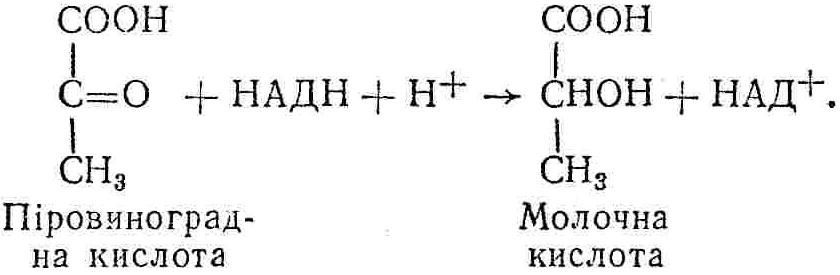 Реакції гліколізу (глікогенолізу) часто поділяють на дві стадії. На першій стадії проходить енергозалежний процес фосфорилювання глюкози та розкладання її на дві тривуглецеві сполуки — фосфотріози. Реакції першої стадії ендоергічні, вони проходять з поглинанням енергії АТФ. Цю стадію перетворення глюкози можна вважати підготовчою. Друга стадія гліколізу (глікогенолізу) забезпечує перетворення фосфотріоз на кінцевий продукт — молочну кислоту. В зв'язку із специфічністю частини реакцій, які відбуваються на цій стадії, вони дістали назву гліколітичної оксидоредукції. В процесі утворення молочної кислоти синтезується також макроергічна сполука АТФ, тобто друга стадія гліколізу проходить з виділенням енергії і є екзергонічним процесом. Як видно з наведених вище рівнянь хімічних реакцій, під час гліколізу з однієї молекули глюкози утворюється дві молекули фосфотріоз. Кожна з них у процесі перетворення на молочну кислоту зумовлює утворення двох молекул АТФ, тобто всього утворюється чотири молекули АТФ. Однак дві з них використовуються для фосфорилювання глюкози (гексокіназна реакція) та утворення фруктозо-1,6-дифосфату (фруктокіназна реакція), тому енергетична ефективність гліколізу становить дві молекули АТФ. Оскільки в макроергічному зв'язку АТФ акумулюється в середньому 42 кДж енергії, то всього під час гліколізу нагромаджується 84 кДж енергії. Експериментально установлено, що внаслідок перетворення глюкози у дві молекули молочної кислоти зміна вільної енергії дорівнює 210 кДж/моль. На основі цього неважко обчислити, що під час гліколізу близько 40 % вивільненої енергії акумулюється в макроергічних зв'язках АТФ, а решта розсіюється у вигляді теплоти. Отже, коефіцієнт корисної дії гліколізу становить 0,35—0,40. Загальну схему гліколізу можна записати так: Під час глікогенолізу також утворюється чотири молекули АТФ, однак на першій стадії цього процесу використовується лише одна молекула АТФ (у фосфофруктокіназній реакції), тому енергетичний ефект глікогенолізу становить 3 молекули АТФ, або 126 кДж/моль. На перший погляд енергетичний ефект глікогенолізу більший, ніж гліколізу. Однак, якщо врахувати, що частина молекул АТФ використовується для синтезу глікогену, стане зрозумілим, що ці два процеси в енергетичному відношенні майже рівноцінні. Для організму і гліколіз, і глікогеноліз не вигідні, оскільки для компенсування енергетичних витрат необхідна велика кількість вуглеводів. Однак як фізіологічні процеси вони досить важливі, оскільки дають змогу забезпечити організм енергією за умов недостатнього постачання тканин киснем. Під час вивчення енергетичних ефектів окремих реакцій анаеробного перетворення вуглеводів було встановлено, що більшість реакцій цього процесу близькі до рівноваги і можуть проходити як у прямому, так і в зворотному напрямках. Це дає змогу організму з одних і тих самих речовин діставати як хімічну енергію, так і сполуки, необхідні для забезпечення протікання різних метаболічних реакцій. Однак три реакції анаеробного перетворення вуглеводів (гексокіназна, фруктокіназна і піруваткіназна) супроводжуються значним зменшенням вільної енергії, тому є практично необоротними. Перебіг цих реакцій у зворотному напрямку потребує подолання певного енергетичного бар'єру та наявності специфічних ферментів. Спиртове бродіння. У нижчих організмів — дріжджових і цвільових грибків, деяких мікроорганізмів — процес анаеробного перетворення вуглеводів завершується утворенням етилового спирту, тому він дістав назву спиртового бродіння: Хімізм цього процесу досить близький до гліколізу, який протікає в тканинах вищих організмів. Усі стадії перетворення глюкози до утворення піровиноградної кислоти включно в обох випадках проходять однаково і каталізуються одними і тими самими ферментними системами. Відмінність між цими процесами виявляється, починаючи з етапу перетворення піровиноградної кислоти. Так, підчас гліколізу і глікогенолізу піровиноградна кислота відновлюється до молочної, а під час спиртового бродіння вона піддається декарбоксилюванню і перетворюється на оцтовий альдегід:  Фермент піруватдекарбоксилаза, що каталізує цю реакцію, містить у вигляді простетичної групи тіамінпірофосфат, який зв'язується з піровиноградною кислотою з утворенням оксіетилтіамінпірофосфату. В процесі перетворення останнього вивільнюється оцтовий альдегід і тіамінпірофосфат. Каталітична активність ферменту забезпечується наявністю іонів Mg2+. Утворений за цією реакцією оцтовий альдегід далі за участю НАД•Н+Н+ відновлюється до кінцевого продукту спиртового бродіння — етилового спирту: 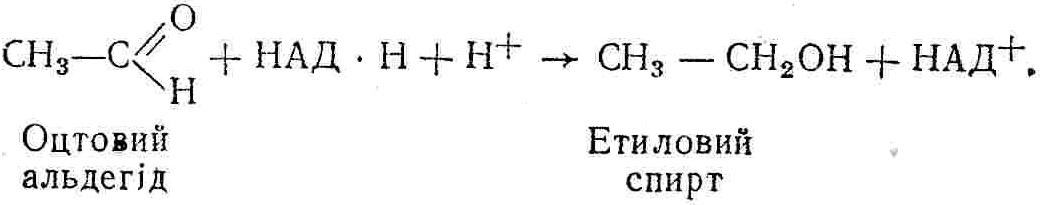 Реакцію каталізує фермент алкогольдегідрогеназа. 4. АЕРОБНЕ ПЕРЕТВОРЕННЯ ВУГЛЕВОДІВ Аеробне й анаеробне перетворення вуглеводів тісно зв'язані між собою. Це насамперед виявляється в тому, що обидва процеси проходять однаково включно до стадії утворення піровиноградної кислоти. В них беруть участь одні й ті самі ферменти та утворюються однакові проміжні продукти. Відмінність між анаеробним і аеробним розкладанням вуглеводів починається з перетворення піровиноградної кислоти. Якщо вуглеводи перетворюються в анаеробних умовах, то піровиноградна кислота, як уже зазначалося, відновлюється до молочної кислоти; при перетворенні вуглеводів в аеробних умовах піровиноградна кислота піддається декарбоксилюванню з утворенням ацетил-КоА, який далі окислюється до кінцевих продуктів — СО2 і Н2О з виділенням значної кількості енергії, що акумулюється в молекулах АТФ. Окислювальне декарбоксилювання піровиноградної кислоти Процес перетворення піровиноградної кислоти до ацетил-КоА дістав назву окислювального декарбоксилювання.Каталізується він складним поліферментним комплексом — піруватдегідрогеназою, який пов'язаний з дією п'яти коферментів: тіамінпірофосфату, ліпоєвої кислоти, коензиму А, НАД і ФАД. Численними дослідженнями доведено, що окислювальне декарбоксилювання піровиноградної кислоти в тканинах організмів людини і тварин проходить лише в аеробних умовах. Загальну схему даного процесу можна подати так: 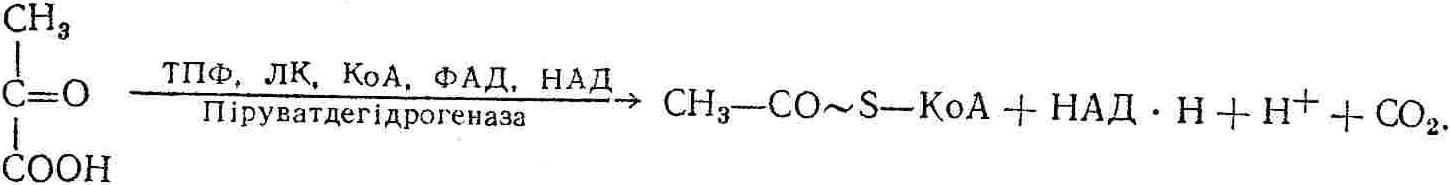 В зв'язку із значним зменшенням вільної енергії ця реакція є необоротною. Вона проходить у декілька стадій. На першій стадії піровиноградна кислота взаємодіє з тіамінпірофосфатом з утворенням проміжної сполуки, де вона перебуває в активному стані. Далі під каталітичною дією піруватдекарбоксилази піровиноградна кислота, що входить до складу тіамінпірофосфату, декарбоксилюється. В результаті цієї реакції утворюється оксіетилтіамінпірофосфат, який під впливом ферменту ліпоїлредуктази ацетилтрансферази, коферментом якого є ліпоєва кислота, розкладається на вільний тіамінпірофосфат і ацетилліпоєву кислоту. Ацетилліпоєва кислота утворюється внаслідок переходу залишку оцтової кислоти від оксіетилтіамінпірофосфату до ліпоєвої кислоти: 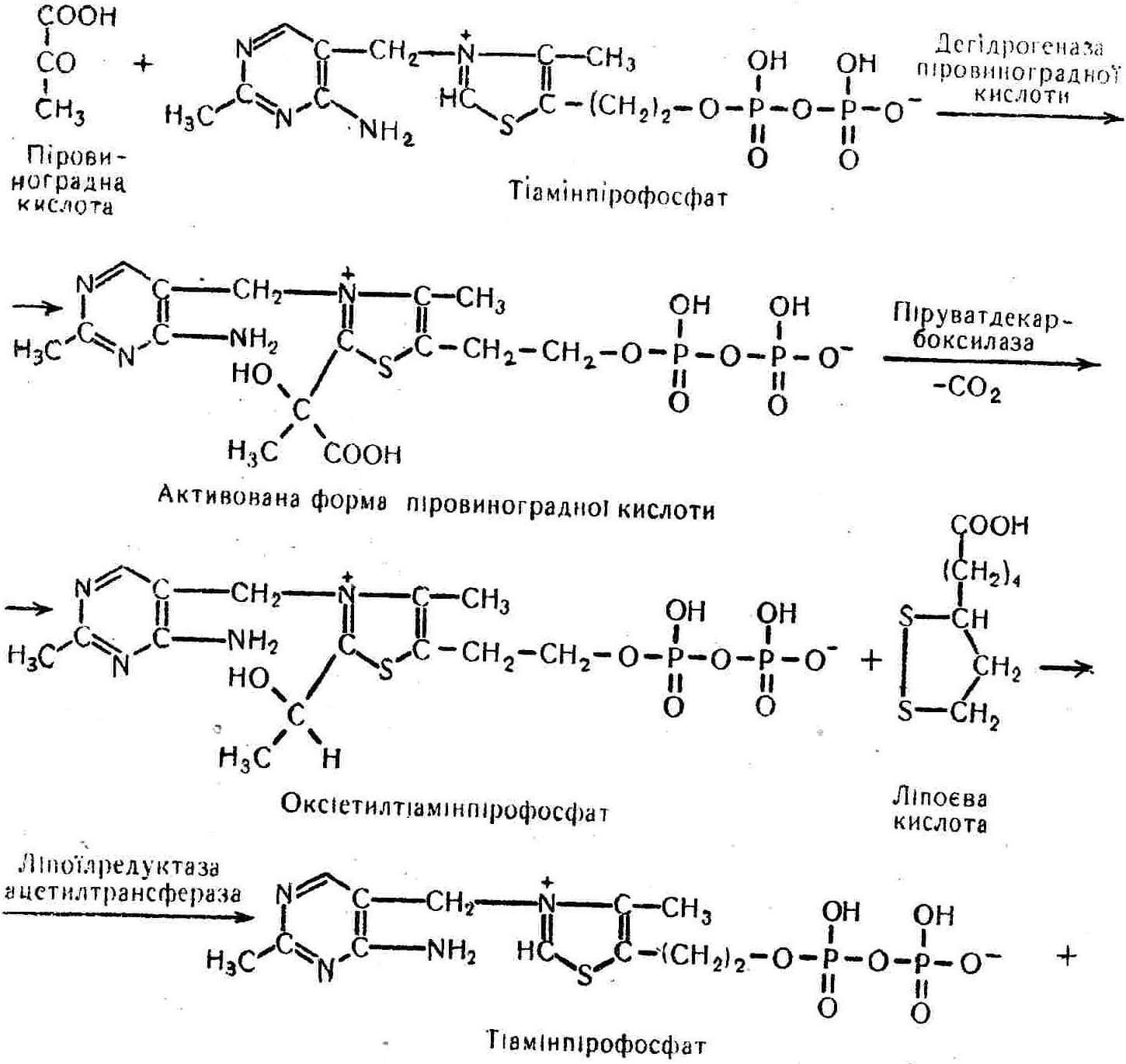 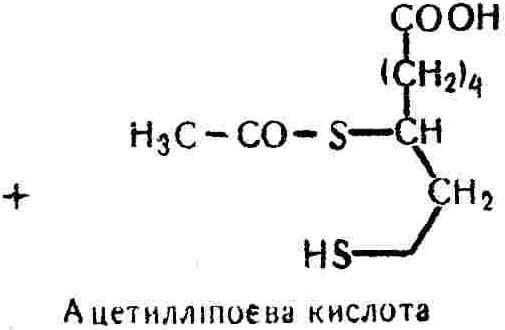 З ацетилліпоєвої кислоти залишок ацетилу далі переноситься на КоА—SН. Продуктами реакції є ацетил-КоА і дигідроліпоєва кислота. Слід підкреслити, що дигідроліпоєва кислота під впливом ФАД здатна переходити у ліпоєву кислоту: 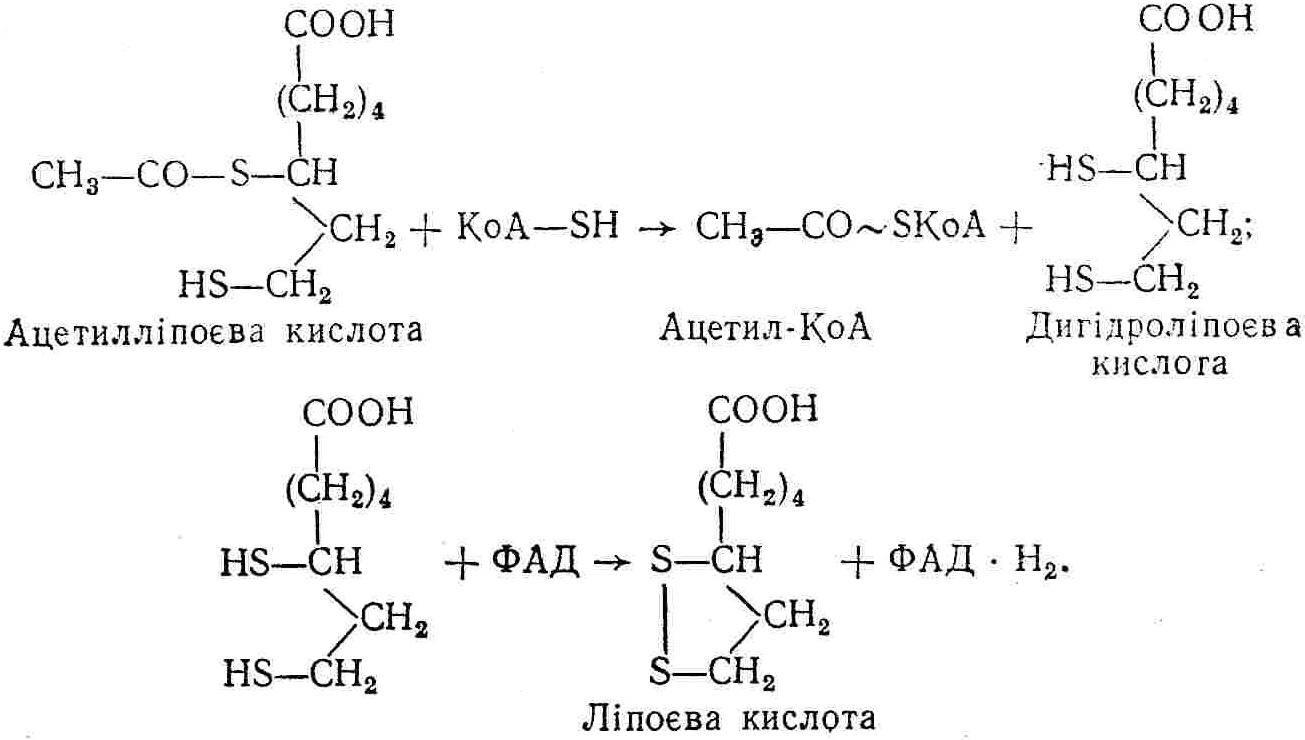 За цих умов відновлений ФАД передає атоми водню на окислений НАД: Відновлений НАД, що утворився, за допомогою цитохромної системи окислюється киснем повітря: В результаті окислення, спряженого з процесами фосфорилювання, синтезується три молекули АТФ, тобто акумулюється 126 кДж енергії. Частина ацетил-КоА, що утворився під час декарбоксилювання піровиноградної кислоти, використовується для синтезу жирів, вуглеводів та інших сполук, а інша його частина вступає в цикл трикарбонових кислот, де окислюється до СО2 і Н2О. При цьому вивільнюється певна кількість енергії. Цикл трикарбонових кислот (цикл Кребса) Цикл трикарбонових кислот є одним з найголовніших шляхів окислення залишків оцтової кислоти до кінцевих продуктів. Він властивий всім тканинам організмів людини і вищих тварин, а також деяким аеробним мікроорганізмам. Цей цикл є заключною стадією окислювального катаболізму не лише вуглеводів, а й інших класів органічних сполук. На першій стадії циклу трикарбонових кислот відбувається так звана цитратсинтетазна реакція (утворення лимонної кислоти) внаслідок взаємодії ацетил-КоА з щавлевооцтовою кислотою. Реакція проходить у кілька стадій і каталізується одним ферментом — цитратсинтетазою. Залежно від стадії, яку каталізує фермент, він виконує роль енолази, синтетази або гідролази. Спочатку протікає реакція енолізації щавлевооцтової кислоти: 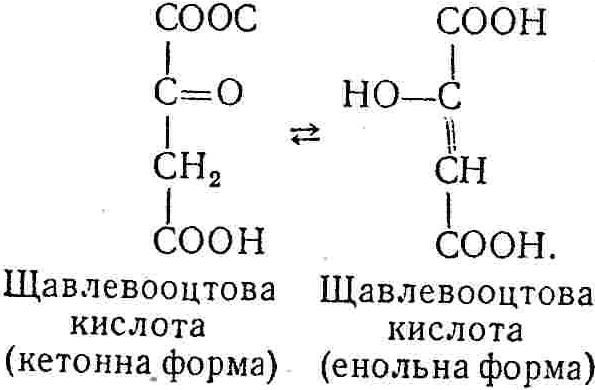 Далі енольна форма щавлевооцтової кислоти взаємодіє з ацетил-КоА внаслідок чого утворюється цитрил-КоА, що гідролізується з утворенням лимонної кислоти: 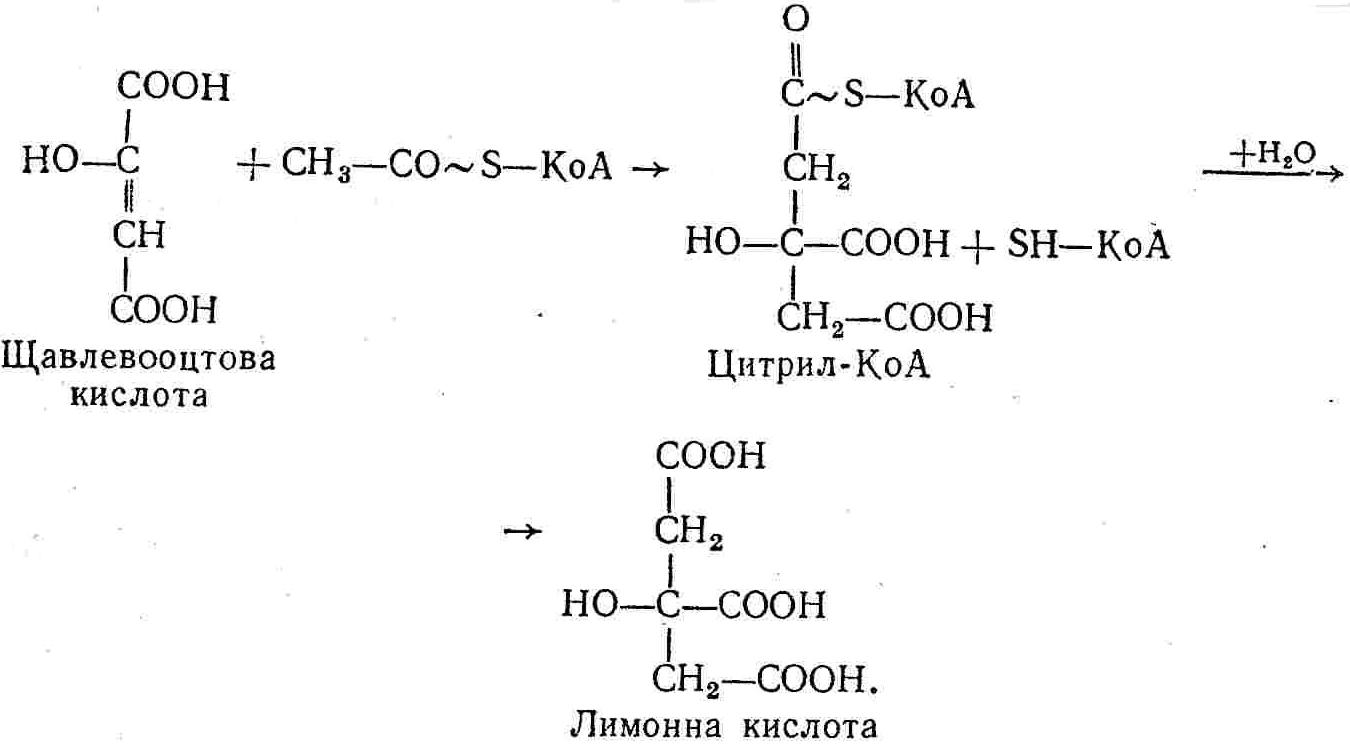 Далі лимонна кислота за участю ферменту аконітатгідратази перетворюється на ізолимонну кислоту: 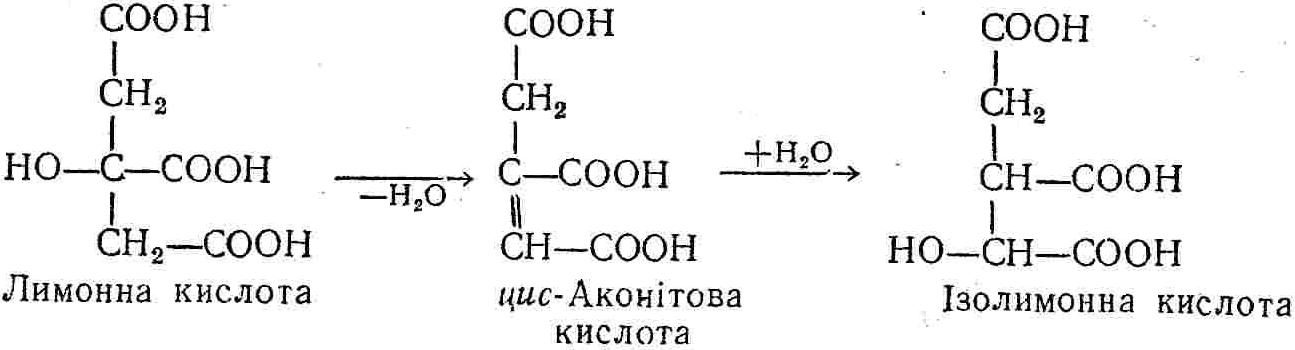 Остання під впливом ферменту ізоцитратдегідрогенази окислюється і перетворюється на щавлевоянтарну кислоту. За цих умов атоми водню приймає НАДФ+: 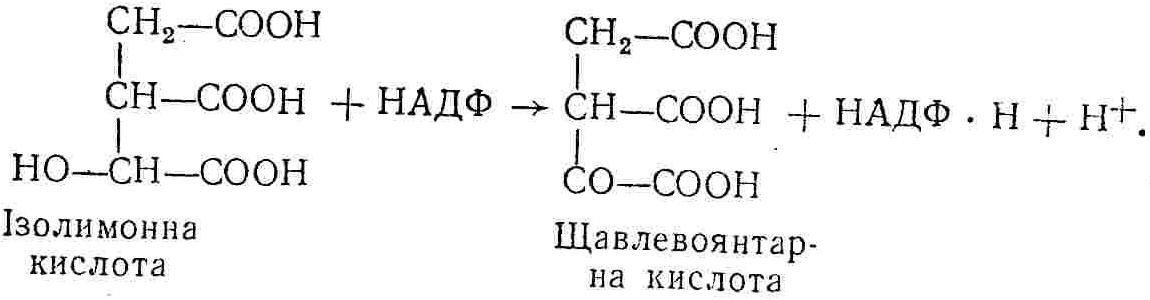 Щавлевоянтарна кислота при каталітичній дії специфічної декарбоксилази декарбоксилюється з утворенням α-кетоглутарової кислоти: 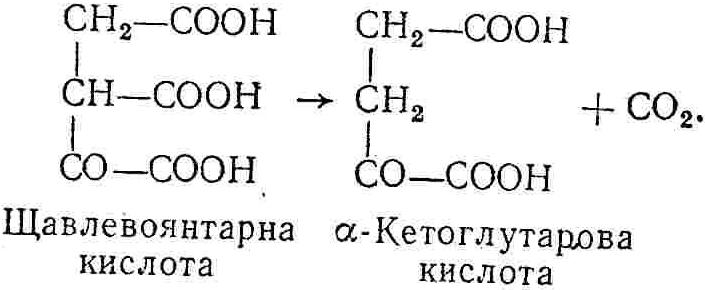 α-Кетоглутарова кислота декарбоксилюється повторно. Реакція відбувається подібно до декарбоксилювання піровиноградної кислоти за участю коферментів НАД+, ліпоєвої кислоти, КоА і ТПФ. Продуктом реакції є сукциніл-КоА: 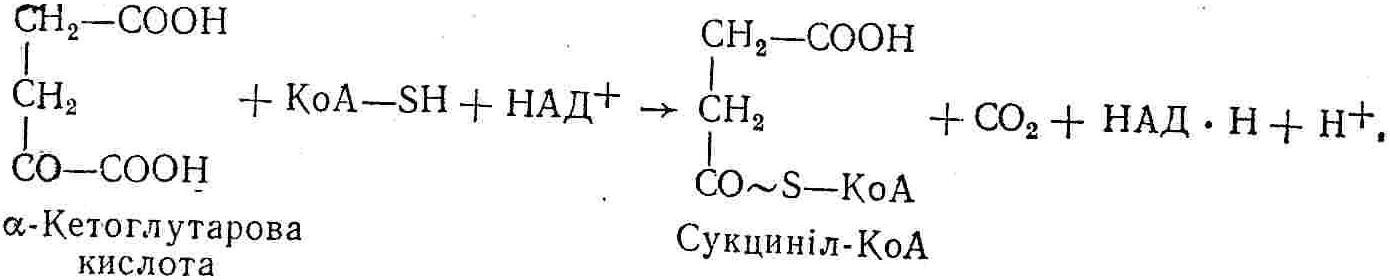 Далі сукциніл-КоА перетворюється на янтарну кислоту. Енергія, акумульована в карбонілтіоефірному зв'язку, переходить у макроергічний зв'язок ГТФ: 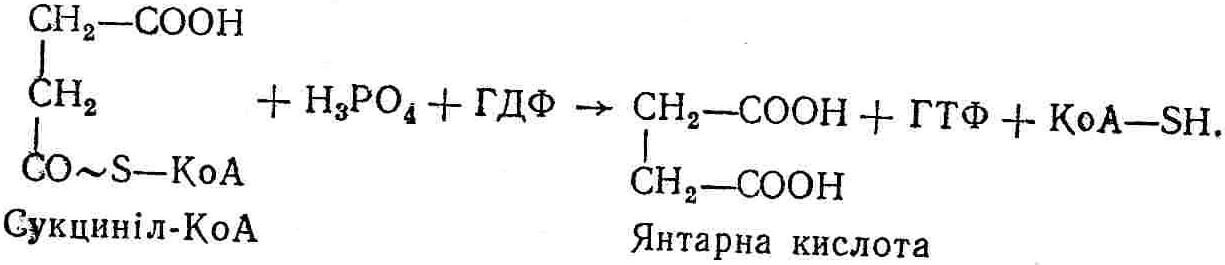 ГТФ за участю ферменту ГТФ-аденілаткінази передає макроергічний зв'язок із залишком фосфорної кислоти на АДФ. При цьому утворюється АТФ: ГТФ + АДФ ГДФ + АТФ. Під впливом ферменту сукцинатдегідрогенази, коферментом якої є ФАД, янтарна кислота окислюється до фумарової кислоти:  Під час гідратації за участю ферменту фумаратгідратази фумарова кислота перетворюється на яблучну кислоту:  Завершується цикл Кребса окисленням яблучної кислоти під впливом ферменту малатдегідрогенази, що містить кофермент НАД+. Продуктом реакції є щавлевооцтова кислота, тобто той субстрат, з якого розпочинався трикарбоновий цикл:  Щавлевооцтова кислота, що утворилась, переходить в енольну форму, вступає в реакцію з новою молекулою ацетил-КоА і далі повторюється весь цикл перетворень. Так, відносно невелика кількість щавлевооцтової кислоти, вступаючи кілька разів у реакцію конденсації з ацетил-КоА, забезпечує окислення значної кількості оцтової кислоти, яка утворюється в процесі обміну не тільки вуглеводів, а й ліпідів і білків. Внаслідок усіх перетворень циклу Кребса оцтова кислота у вигляді ацетил-КоА розкладається на СО2 і Н2О: Енергія, що вивільнюється під час окислення оцтової кислоти, акумулюється в макроергічних зв'язках АТФ. Як видно з рівнянь реакцій, у циклі Кребса утворюється чотири молекули відновлених форм коферментів: одна молекула НАДФ•Н+Н+, дві молекули НАД•Н+Н+ і одна молекула ФАД•Н2. При цьому доведено, що під час окислення однієї молекули НАД•Н+Н+ або НАДФ•Н+Н+ шляхом відщеплення атома водню в ланцюгу дихальних ферментів синтезується три молекули АТФ. Внаслідок окислення ФАД•Н2 утворюється дві молекули АТФ. Отже, всього під час окислення відновлених форм коферментів утворюється 3•3 + 2 = 11 молекул АТФ. Крім того, одна молекула АТФ утворюється на рівні субстрату під час перетворення сукциніл-КоА в янтарну кислоту. Отже, в процесі перетворення однієї молекули ацетил-КоА в циклі трикарбонових кислот синтезується 11 + 1 = 11 молекул АТФ. Разом з тим три молекули АТФ синтезуються внаслідок окислення однієї молекули НАД•Н+Н+, яка утворюється в процесі перетворення піровиноградної кислоти до ацетил-КоА. Всього в процесі перетворення однієї молекули піровиноградної кислоти до ацетил-КоА і останнього до СО2 і Н2О синтезується 12 + 3 = 15 молекул АТФ. Оскільки з однієї молекули глюкози утворюється дві молекули піровиноградної кислоти, то всього утворюється 15•2 = 30 молекул АТФ. Крім того, 6 молекул АТФ утворюється внаслідок окислення двох молекул НАД•Н+Н+, які вивільнюються під час гліколітичної оксиредукції, і дві молекули АТФ синтезується в процесі гліколізу. Тому загальний енергетичний ефект аеробного розкладання однієї молекули глюкози до кінцевих продуктів — СО2 і Н2О — становить 30 + 6 + 2 = 38 молекул АТФ. Оскільки в одній молекулі АТФ зосереджено 42 кДж енергії, то всього під час аеробного перетворення однієї молекули глюкози акумулюється 1596 кДж енергії. Перетворення однієї молекули глюкози за анаеробних умов, як зазначалось раніше, дає лише дві молекули АТФ, тобто в макроергічних зв'язках її акумульовано 2•42 = 84 кДж енергії. Отже, основним джерелом енергії для організму є аеробне окислення органічних сполук. |