Аденилатциклазная система. Аденилатциклаза сообщение. Аденилатциклаза
 Скачать 98.22 Kb. Скачать 98.22 Kb.
|

|
Аденилатцикла́за (АЦ, англ. AC, adenylate cyclase, adenylyl cyclase— фермент[2], который катализирует превращение АТФ в 3',5'-цАМФ (циклическую форму АМФ) с образованием пирофосфата . В процессе передачи сигнала аденилатциклаза может быть активирована связанными с плазматической мембраной рецепторами, связанными с G-белками (GPCR), которые передают гормональные и иные стимулы в клетку. Активация аденилатциклазы приводит к образованию цАМФ, действующего как вторичный посредник. цАМФ взаимодействует с протеинкиназой А, ионными каналами, связанными с циклическими нуклеотидами, и регулирует их функции. ЦиклоАМФ является наиболее популярным мессенджером Аденилатциклазная система При участии аденилатциклазной системы реализуются эффекты сотни различных по своей природе сигнальных молекул – гормонов и нейромедиаторов. Функционирование системы трансмембранной передачи сигналов обеспечивают 5 белков. Интегральные белки цитоплазматической мембраны: Rs-рецептор активатора аденилатциклазной системы и Ri-рецептор ингибитора аденилатциклазной системы Фермент аденилатциклаза «заякоренныве белки»: Gs-ГТФ-связывающий белок, состоит из αs, β,γ -субъединиц и Gj-ГТФ-связывающий белок, состоит из αi, β,γ -субъединиц и одного цитозольного фермента протеинкиназы А (ПКА). Последовательность событий, приводящих к активации аденилатциклазы:Рецептор имеет два центра связывания: для гормона на наружной поверхности мембраны и для G-белка на внутренней поверхности мембраны. Взаимодействие активатора аденилатциклазной системы, например гормона с рецептором (Rs), приводит к изменению конформации рецептора Увеличивается сродство рецептора к Gs-белку. Образуется комплекс гормон-рецептор. Присоединение комплекса гормон-рецептор к Gs-ГДФ снижает сродство α-субъединицы Gs-белка к ГДФ и увеличивает сродство к ГТФ. В активном центре αs-субъединицы ГДФ замещается на ГТФ; Это вызывает изменение конформации αs-субъединицы и снижение сродства к βγ-субъединицам Отделившаяся αs-ГТФ субъединица латерально перемещается в липидном слое мембраны к центру связывания фермента аденилатциклазы Взаимодействие αs- ГТФ субъединицы с аденилатциклазой приводит к изменению конформации фермента, его активации и увеличению скорости образования цАМФ из АТФ; В клетке повышается концентрация цАМФ – вторичного посредника гормонального сигнала Молекулы цАМФ могут обратимо соединяться с регуляторными субъединицами протеинкиназы А, которая состоит из 2 регуляторных и 2 каталитических субъединиц Присоединение цАМФ к регуляторным субъединицам вызывает диссоциацию комплекса, каталитические субъединицы отделяются и становятся активными Активная протеинкиназа А с помощью АТФ фосфорилирует специфические белки по серину и треонину, в результате изменяются конформация и активность фосфорилированных белков, а это приводит к изменению скорости и направления регулируемых ими процессов в клетке. Каскадный механизм усиления и подавления сигнала. Передача сигнала от мембранного рецептора через G-белок на фермент аденилатциклазу служит примером каскадной системы усиления этого сигнала. Одна молекула, активирующая рецептор, может "включать" несколько G-белков, и затем каждый активирует несколько молекул аденилатциклазы с образованием тысяч молекул цАМФ. На этом этапе сигнал усиливается в 102-103 раз. Образующийся цАМФ "включают" другой фермент - протеинкиназу А, усиливая сигнал ещё в 1000 раз. Фосфорилирование ферментов протеинкиназой А ещё больше усиливает сигнал, в результате суммарное усиление равно 106-107 раз. Таким образом, по механизму каскадного усиления одна молекула регулятора способна изменить активность миллионов других молекул. Инозитолфосфатная система Инозитолфосфатная система включает 3 мембранных белка: R- рецептор активатора инозитолфосфатной системы – интегральный белок Фосфолипаза С - поверхностный белок Gplc – ГТФ-связывающий белок активирует фосфолипазу С – заякоренный белок Работу системы обеспечивают белок кальмодулин, фермент протеин киназа С, регулируемые Са2+ -каналы мембраны ЭПР, Са2+ -АТФаза клеточной и митохондриальной мембран 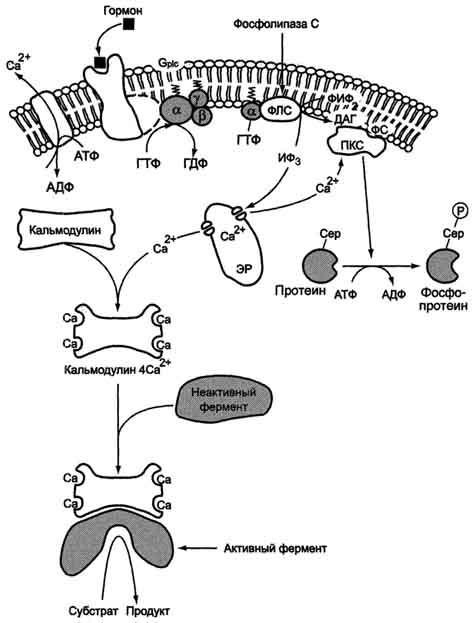 Последовательность событий, приводящих к активации фосфолипазы С: связывание сигнальной молекулы, например гормона с рецептором (R), вызывает изменение конформации и увеличение сродства к Gр1с-белку. образование комплекса гормон-рецептор Gрlс-ГДФ приводит к снижению сродства αрlс-протомера Gрlс-белка к ГДФ и увеличению сродства к ГТФ. В активном центре αрlс-субъединицы ГДФ заменяется на ГТФ. это вызывает изменение конформации αрlс-субъединицы и она отделяется от βγ- субъединицы отделившаяся α- ГТФ субъединица латерально перемещается по мембране к центру связывания фермента фосфолипазы С. взаимодействие α- ГТФ субъединицы с фосфолиггазой С изменят конформацию и активность фермента, увеличивается скорость гидролиза фосфолипида клеточной мембраны фосфатидилинозитол-4,5-бисфосфата (ФИФ2). в ходе гидролиза образуется и выходит в цитозоль гидрофильное вещество инозитол-1,4,5-трифосфат (ИФ3). Другой продукт реакции диацилглицерол (ДАГ) остаётся в мембране и участвует в активации фермента протеинкиназы С (ПКС). инозитол-1,4,5-трифосфат (ИФ3) связывается специфическими центрами Са2+-канала мембраны ЭР, это приводит к изменению конформации белка и открытию Са2+-канала. Са2+ по градиенту концентрации поступает в цитозоль. В отсутствие в цитозоле 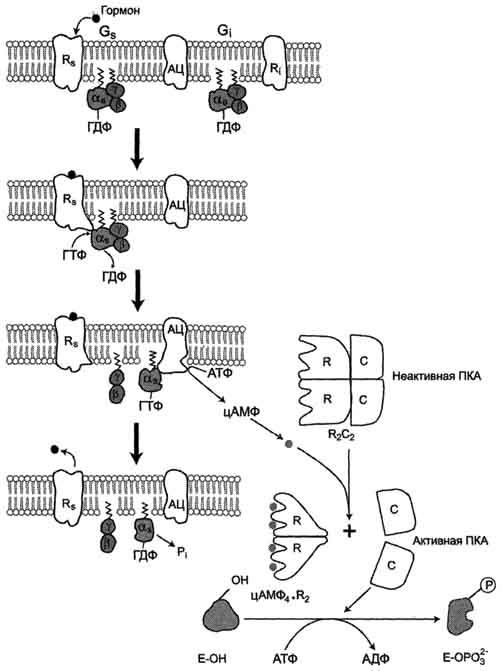 Циклический АМФ (циклоАМФ, цАМФ) образуется в клетке, когда действуют гормоны гипофиза (ТТГ, ЛГ, МСГ, ФСГ. АКТГ), кальцитонин, соматостатин, глюкагон, паратгормон, адреналин (через α2- и β-адренорецепторы), вазопрессин (через V2-рецепторы). Механизм наработки цАМФ связан с активацией фермента аденилатциклазы и называется аденилатциклазный механизм:. Этапы передачи сигнала выглядят следующим образом: 1. Взаимодействие лиганда с рецептором приводит к изменению конформации последнего. Это изменение передается на G-белок (GTP, ГТФ-зависимый), который состоит из трех субъединиц (α, β и γ), α-субъединица связана с ГДФ. В составе G-белков α-субъединицы бывают двух типов по отношению к аденилатциклазе: активирующие αs и ингибирующие αi. Например, в случае действия адреналина с активирующей субъединицей αs взаимодействуют β-адренорецепторы, с ингибирующей – α2-адренорецепторы. 2. В результате взаимодействия с рецептором β- и γ-субъединицы отщепляются, одновременно на α-субъединице ГДФ заменяется на ГТФ. 3. Активированная таким образом αs-субъединица стимулирует аденилатциклазу, которая начинает синтез цАМФ. Если в действо была вовлечена αi-субъединица, то она ингибирует аденилатциклазу, все останавливается. 4. Циклический АМФ (циклоАМФ, цАМФ) – вторичный мессенджер – в свою очередь связывается с регуляторными (R) субъединицами протеинкиназы А и вызывает их диссоциацию от каталитических. В результате каталитические (C) субъединицы становятся активными. Протеинкиназа А (ПК А) фосфорилирует ряд ферментов, среди которых киназа фосфорилазы гликогена, гликогенсинтаза, гормончувствительная липаза, транскрипционный фактор CREB (cAMP-response element-binding protein). 5. Наработка цАМФ продолжается некоторое время, пока α-субъединица, которая является ГТФ-азой, отщепляет фосфат от ГТФ. 6. Как только ГТФ превратился в ГДФ, то α-субъединица инактивируется, теряет свое влияние на аденилатциклазу, обратно соединяется с β- и γ-субъединицами. 7. Все возвращается в исходное положение Гормон отрывается от рецептора еще раньше: если концентрация гормона в крови велика, то следующая его молекула присоединится к рецептору через малый промежуток времени и повторный запуск АЦ-механизма произойдет быстро – в клетке активируются соответствующие процессы. если гормона в крови мало – для клетки наступает некоторая пауза, изменения метаболизма нет. Остановка АЦ-механизмаОчевидно, что находящийся в цитозоле цАМФ должен своеременно удаляться, чтобы гормональный сигнал не оказывал избыточное воздействие. Для ликвидации цАМФ (и циклоГМФ) в клетках присутствуют различные фосфодиэстеразы, "открывающие" созданный фосфатом цикл. 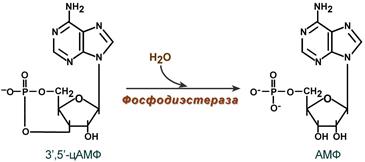 Снижение активности цАМФ-фосфодиэстеразы вызывается глюкокортикоидами, что усиливает действие адреналина при стрессе, и кофеином, что обусловливает его адреналиноподобные эффекты, а именно гипергликемию, ино- и хронотропный эффект на сердце, расширение сосудов скелетных мышц и головного мозга. Активация фосфодиэстеразы вызывается инсулином, также быстро и обратимо обеспечивается ионами Cа2+ либо непосредственно, либо через гуанилатциклазный механизм. |