Ответы на билеты биохимия 2012. Белок это последовательность ак, связанных друг с другом пептидными связями
 Скачать 5.15 Mb. Скачать 5.15 Mb.
|

|
Задача регуляторных систем — сохранение гомеостаза. Обязательным для регуляции является наличие прямых и обратных связей между регулятором и регулируемым объектом. С помощью этих связей осуществляется интеграция и координация. Интеграция — это объединение элементов системы в единое целое. Координация (соподчинение) — это подчинение менее важных элементов системы более важным элементам. Интеграция и координация — это две стороны процесса регуляции. Различают:1.Внутриклеточную регуляцию (ауторегуляцию).2.Дистантную регуляцию (межклеточную). Механизмы клеточной ауторегуляции: 1.Компартментализация (мембранный механизм).Роль мембран состоит в следующем:а)мембраны делят клетки на отсеки и в каждом из них осуществляются свои процессы;б)мембраны обеспечивают активный транспорт и регулируют потоки молекул в клетке и из клетки;в)в мембраны встроены ферменты;г)мембраны защищают клетку от внешних воздействий. Воздействием на функции мембран клетка может регулировать тот или иной процесс. 2.Изменение активности ферментов 3.Изменение количества ферментов. Классификация межклеточных регуляторов 1.Анатомо-физиологическая: а)Гормоны — межклеточные регуляторы, доставляемые к клеткам-мишеням током крови. Вырабатываются в эндокринныхжелезах или рассеянных железистых клетках. б)Нейрогормоны вырабатываются нервными клетками и выделяются в синаптическую щель, то есть в непосредственной близости от клетки-мишени. Нейрогормоны делятся на медиаторы и модуляторы. Медиаторы обладают непосредственным пусковым эффектом. Модуляторы изменяют эффект медиаторов. Примерами медиаторов являются ацетилхолин и норадреналин; модуляторов — у-аминомасляная кислота, дофамин. в) Локальные гормоны - это межклеточные регуляторы, действующие на близлежащие к месту их синтеза клетки. Пример: гормоны,-производные жирных кислот. 2.Классификация по широте действия: а) Гормоны универсального действия действуют на все ткани организма (например, катехоламины, глюкокортикостероиды). б)Гормоны направленного действия действуют на определенные органы-мишени (например, АКТГ действует на кору надпочечников). 3.Классификация по химическому строению: а)Белково-пептидные гормоны: Олигопептиды (кинины, АДГ), Полипептиды (АКТГ, глюкагон), Белки (СТГ, ТТГ, ПТ} б 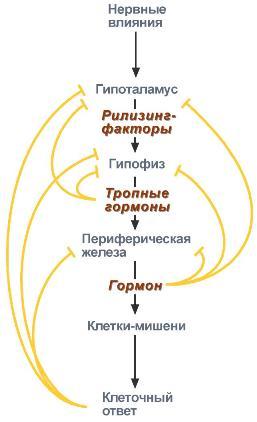 )Производные аминокислот: Катехоламины и йодтиронины — образуются из тирозина;Ацетилхолин — образуется из серина; Серотонин, триптамин, мелатонин — образуются из триптофана. )Производные аминокислот: Катехоламины и йодтиронины — образуются из тирозина;Ацетилхолин — образуется из серина; Серотонин, триптамин, мелатонин — образуются из триптофана.в)Липидные гормоны: стероидные гормоны (гормоны коры надпочечников и половые гормоны); производные полиненасыщенных жирных кислот (простагландины, тромбоксаны, лейкотриены). У многоклеточных существ всегда стоит задача обеспечения взаимосвязи разных органов и баланса их активности. Поэтому большинство гормональных систем взаимосвязаны между собой и регулируются в соответствии с иерархической лестницей. У высших животных верхнюю часть лестницы занимает система гормонов гипоталамуса, контролируемая центральной нервной системой. Сигналы, получаемые от органов, принимаются и обрабатываются, после чего клетки гипоталамуса отвечают при помощи специфических сигнальных молекул – рилизинг-факторов. На стимулирующие или тормозящие стимулы из ЦНС секретируются стимулирующие или ингибирующие рилизинг-факторы, которые носят название либерины или с 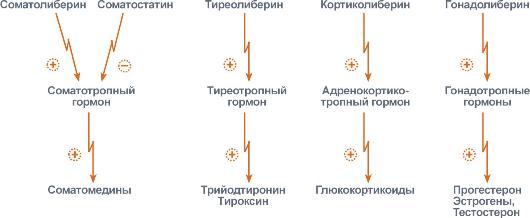 татины соответственно. Эти нейрогормоны с кровотоком достигают аденогипофиза, где стимулируют (либерины) или ингибируют (статины) биосинтез и секрецию тропных гормонов. татины соответственно. Эти нейрогормоны с кровотоком достигают аденогипофиза, где стимулируют (либерины) или ингибируют (статины) биосинтез и секрецию тропных гормонов.Тропные гормоны воздействуют на периферические железы, стимулируя выделение соответствующих периферических гормонов. К подобным системам относятся группы гормонов тиреоидной функции, глюкокортикоидной функции и профиль половых гормонов. Регуляция этих систем осуществляется по принципу обратной отрицательной связи, т.е. накопление гормонов периферических желез тормозит секрецию рилизинг-факторов гипоталамуса и тропных гормонов гипофиза. Наиболее клинически значимо это проявляется в отношении регуляции стероидных гормонов. Подавляющее действие на активность эндокринных желез может оказывать и результат ответа клеток-мишеней. Существуют эндокринные железы для которых отсутствует регуляция тропными гормонами – паращитовидная железа, мозговое вещество надпочечников, ренин-альдостероновая система и поджелудочная железа. Они контролируются нервными влияниями или концентрацией определенных веществ в крови. Семейство гипоталамических гормонов – рилизинг-факторов – включает вещества, как правило небольшие пептиды, образующиеся в ядрах гипоталамуса. Их функция – регуляция секреции гормонов аденогипофиза: стимулирование – либерины и подавление – статины. Доказано существование семи либеринов и трех статинов. Тиреолиберин – является трипептидом, стимулирует секрецию тиреотропного гормона и пролактина, также проявляет свойства антидепрессанта. Кортиколиберин – полипептид из 41 аминокислоты, стимулирует секрецию АКТГ и ?-эндорфина, широко влияет на деятельность нервной, эндокринной, репродуктивной, сердечно-сосудистой и иммунной систем. Гонадолиберин (люлиберин) – пептид из 10 аминокислот, стимулирует высвобождение лютеинизирующего и фолликулостимулирующего гормонов. Гонадолиберин присутствует также в гипоталамусе, участвуя в центральной регуляции полового поведения. Фоллиберин – стимулирует высвобождение фолликулостимулирующего гормона. Пролактолиберин – стимулирует секрецию лактотропного гормона. Пролактостатин – предполагается, что он является дофамином. Снижает синтез и секрецию лактотропного гормона. Соматолиберин состоит из 44 аминокислот и повышает синтез и секрецию гормона роста. Соматостатин – пептид из 12 аминокислот, ингибирующий секрецию ТТГ, пролактина, АКТГ и СТГ из гипофиза. Он образуется также в островках поджелудочной железы и контролирует высвобождение глюкагона и инсулина, а также гормонов желудочно-кишечного тракта. Меланостимулирующий фактор, пентапептид, оказывает стимулирующее действие на синтез меланотропного гормона. Меланостатин, может быть как три-, так и пентапептидом, обладает антиопиоидным эффектом и активностью в поведенческих реакциях. Кроме рилизинг-гормонов в гипоталамусе синтезируются также вазопрессин (антидиуретический гормон) и окситоцин. 48. Понятие о рецепторах. Механизм действия гормонов через внутриклеточные рецепторы и рецепторы плазматических мембран и вторые посредники (общая характеристика). Рецепция и эффект гормона на органы-мишени является основным звеном эндокринной регуляции. Способность гормона к передаче регуляторного сигнала обусловлена наличием в клетках-мишенях специфических рецепторов. Рецепторы в большинстве случаев – белки, преимущественно гликопротеиды, имеющие специфическое фосфолипидное микроокружение. Связывание гормона с рецептором определяется законом действующих масс по кинетике Михаэлиса. При рецепции возможно проявление положительного или отрицательного кооперативных эффектов, когда ассоциация первых молекул гормона с рецептором облегчает или затрудняет связывание последующих. Рецепторный аппарат обеспечивает избирательный прием гормонального сигнала и инициацию специфического эффекта в клетке. Локализация рецепторов в определенной мере обуславливает тип действия гормона. Выделяют несколько групп рецепторов: 1) Поверхностные: при взаимодействии с гормоном меняют конформацию мембран, стимулируя перенос ионов или субстратов в клетку (инсулин, ацетилхолин). 2). Трансмембранные: имеют контактный участок на поверхности и внутримембранную эффекторную часть, связанную с аденилат- или гуанилатциклазой. Образование внутриклеточных мессенджеров – цАМФ и цГМФ – стимулирует специфические протеинкиназы, влияющие на синтез белка, активность ферментов и т.д. (полипептиды, амины). 3) Цитоплазматические: связываются с гормоном и в виде активного комплекса поступают в ядро, где контактируют с акцептором, приводя к усилению синтеза РНК и белка (стероиды). 4) Ядерные: существуют в виде комплекса негистонового белка и хроматина. Контакт с гормоном напрямую включает механизм его действия (гормоны щитовидной железы). Виды мембраносвязанных рецепторов 1 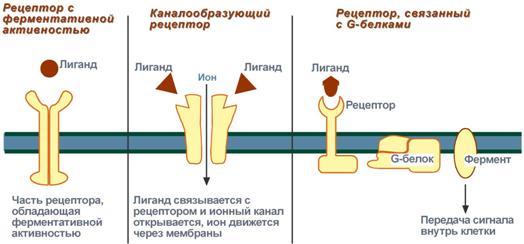 . Рецепторы, обладающие каталитической активностью – при взаимодействии лиганда с рецептором активируется внутриклеточная часть (домен) рецептора, имеющий тирозинкиназную или тирозинфосфатазную или гуанилатциклазную активность. По этому механизму действуют СТГ, инсулин, пролактин, интерлейкины, ростовые факторы, интерфероны α, β, γ. . Рецепторы, обладающие каталитической активностью – при взаимодействии лиганда с рецептором активируется внутриклеточная часть (домен) рецептора, имеющий тирозинкиназную или тирозинфосфатазную или гуанилатциклазную активность. По этому механизму действуют СТГ, инсулин, пролактин, интерлейкины, ростовые факторы, интерфероны α, β, γ.2 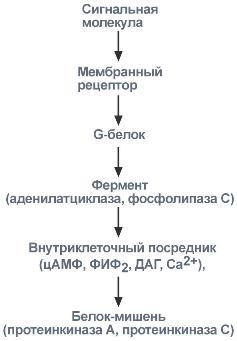 . Каналообразующие рецепторы – присоединение лиганда к рецептору вызывает открытие ионного канала на мембране. Таким образом действуют нейромедиаторы (ацетилхолин, глицин, ГАМК, серотонин, гистамин, глутамат); . Каналообразующие рецепторы – присоединение лиганда к рецептору вызывает открытие ионного канала на мембране. Таким образом действуют нейромедиаторы (ацетилхолин, глицин, ГАМК, серотонин, гистамин, глутамат);3. Рецепторы, связанные с G-белками – передача сигнала от гормона происходит при посредстве G-белка. G-белок влияет на ферменты, образующие вторичные мессенджеры (посредники ). Последние передают сигнал на внутриклеточные белки. Большинство гормонов действуют по данному механизму. К третьему виду относятся аденилатциклазный и кальций-фосфолипидный механизмы. Величина эффекта гормона зависит от концентрации гормонального рецептора, поступающего к клеткам-мишеням, от числа специфических рецепторов, степени их сродства и избирательности к гормону. На величину эффекта может влиять действие других гормонов, как антагонистическое (инсулин и глюкокортикоиды разнонаправленно действуют на поступление глюкозы в клетку), так и потенцирующее (глюкокортикоиды усиливают влияние катехоламинов на сердце и мозг). Изучение функционирования рецепторного аппарата актуально в клинике, особенно при сахарном диабете, вызванном рецепторной инсулинорезистентностью, при синдроме тестикулярной феминизации или определении гормон-чувствительных опухолей молочной железы. Гормоны оказывают свое действие, связываясь со специфическими рецепторами, располагающимися либо на поверхности мембраны клетки, либо в цитозоле. Связывание с рецепторами - обязательный этап в действии гормона. Белково-пептидные гормоны и гормоны-производные аминокислот являются гидрофильными веществами и проникновение их через плазматическую мембрану, состоящую из липидного бислоя, затруднено или невозможно. Рецепторы таких гормонов находятся на наружной поверхности плазматической мембраны. Гормоны связываются с рецепторными белками тех участков мембран клеток-мишеней, которые соприкасаются с кровью, что в свою очередь активирует ферментную систему, отвечающую за образование вторичного (внутриклеточного) посредника. Рецепторы, находящиеся на клеточной мембране. Гормоны (первичные посредники), связываясь с рецепторами на поверхности клеточной мембраны, образуют комплекс гормон-рецептор, который трансформирует сигнал первичного посредника в изменение концентрации особых молекул внутри клетки - вторичных посредников. Вторичными посредниками могут быть следующие молекулы: цАМФ, цГМФ, ИФ3, ДАТ, Са2+, NO. В этом случае гормон не проникает в клетку, а взаимодействует с рецептором на поверхности мембраны. Далее возможны два варианта событий: 1.Первый вариант — с рецептором связан фермент, который из специфического субстрата образует второй посредник. Второй посредник далее связывается со своим рецептором в клетке. Чаще всего рецептором посредника является протеинкиназа, которая за счет фосфата АТФ, фосфорилирует белки. В результате изменяются их свойства, возникает биохимический и физиологический эффект. 2.Второй вариант — рецептор связан не с ферментом мембраны, а с ионным каналом. При связывании гормона с рецептором, канал открывается, ион поступает в клетку и выполняет функции второго посредника. Хорошо изученными вторыми посредниками являются циклические нуклеотиды (цАМФ, цГМФ) и Са2+. мембранный механизм - когда гормон связывается с рецептором на поверхности наружной мембраны клетки-мишени; Гормоны обладающие мембранным механизмом действия: 1) все белковые и пептидные гормоны, а также амины (адреналин, норадреналин). Гормоны оказывают влияние на клетки-мишени. Клетки-мишени - это клетки, которые специфически взаимодействуют с гормонами с помощью специальных белков-рецепторов. Эти белки-рецепторы располагаются на наружной мембране клетки, или в цитоплазме, или на ядерной мембране и на других органеллах клетки. Биохимические механизмы передачи сигнала от гормона в клетку-мишень. Любой белок-рецептор состоит, минимум из двух доменов (участков), которые обеспечивают выполнение двух функций: 1) узнавание гормона; 2) преобразование и передачу полученного сигнала в клетку. Один из доменов белка-рецептора имеет в своем составе участок, комплементарный какой-то части сигнальной молекулы. Процесс связывания рецептора с сигнальной молекулой похож на процесс образования фермент-субстратного комплекса и может определяется величиной константы сродства. Между молекулой гормона и рецептором формируются электростатические и гидрофобные взаимодействия. При связывании рецептора с гормоном происходят конформационные изменения белка-рецептора и комплекс сигнальной молекулы с белком-рецептором активируется. Механизмы действия гормонов на клетки-мишени В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных - рецепторы находятся в наружной мембране. Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют вторыми посредниками. Молекулы гормонов очень разнообразны по форме, а "вторые посредники" - нет. Надежность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору. Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов? Э 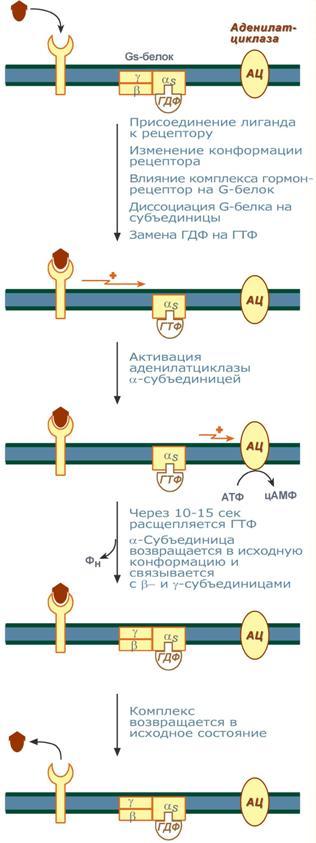 то циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок - кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы - ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях. Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия: то циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок - кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы - ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях. Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия:1) аденилатциклазная (или гуанилатциклазная) системы;2) фосфоинозитидный механизм. Аденилатциклазная система. Циклический АМФ (циклоАМФ, цАМФ) образуется в клетке, когда действуют гормоны гипофиза (ТТГ, ЛГ, МСГ, ФСГ. АКТГ), кальцитонин, соматостатин, глюкагон, паратгормон, адреналин (через α2- и β-адренорецепторы), вазопрессин (через V2-рецепторы). Этапы передачи сигнала: 1) Взаимодействие лиганда с рецептором приводит к изменению конформации последнего. 2)Это изменение передается на G-белок, который состоит из трех субъединиц (α, β и γ), α-субъединица связана с ГДФ.В составе G-белков α-субъединицы бывают 2х типов по отношению к аденилатциклазе: активирующие αS и ингибирующие αI. 3)В результате взаимодействия с рецептором β- и γ-субъединицы отщепляются, одновременно на α-субъединице ГДФ заменяется на ГТФ. 4)Активированная таким образом αS-субъединица стимулирует аденилатциклазу, которая начинает синтез цАМФ. Если в действо была вовлечена αI-субъединица – она ингибирует аденилатциклазу, все останавливается. 5) Циклический АМФ – вторичный мессенджер – в свою очередь, взаимодействует с протеинкиназой А и активирует ее. Протеинкиназа А фосфорилирует ряд ферментов, среди которых киназа фосфорилазы гликогена, гликогенсинтаза, ТАГ-липаза. 6)Наработка цАМФ продолжается некоторое время, пока α-субъединица, которая является ГТФ-азой, отщепляет фосфат от ГТФ. 7 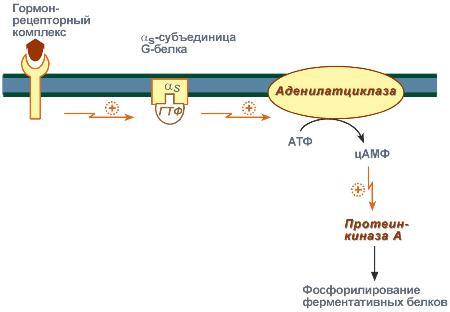 ) Как только ГТФ превратился в ГДФ, то α-субъединица инактивируется, теряет свое влияние на аденилатциклазу, обратно соединяется с β- и γ-субъединицами. Все возвращается в исходное положение. ) Как только ГТФ превратился в ГДФ, то α-субъединица инактивируется, теряет свое влияние на аденилатциклазу, обратно соединяется с β- и γ-субъединицами. Все возвращается в исходное положение.8)Гормон отрывается от рецептора еще раньше: - если концентрация гормона в крови велика, то следующая его молекула присоединится к рецептору через малый промежуток времени и повторный запуск АЦ-механизма произойдет быстро – в клетке активируются соответствующие процессы. - если гормона в крови мало – для клетки наступает некоторая пауза, изменения метаболизма нет. Механизм наработки цАМФ связан с активацией фермента аденилатциклазы и называется аденилатциклазный механизм. Гормоны, для которых циклический АМФ служит в качестве вторичного медиатора: Гормон Мишень: орган/ткань Типичное действие Адренокортикотропный гормон Кора надпочечников Продуцирование кортизола Кальцитонин Кости Концентрация кальция в сыворотке крови Катехоламины (b-адренергические) Сердце ЧСС, сократимость миокарда Хорионический гонадотропин Яичники, семенники Продуцирование половых гормонов Фолликулостимулирующий гормон Яичники, семенники Гаметогенез Глюкагон Печень Гликогенолиз, высвобождение глюкозы Лютеинизирующий гормон Яичники, семенники Продуцирование половых гормонов Рилизинг-фактор лютеинизирующего гормона Гипофиз Высв-е лютеинизирующего гормона Меланоцитстимулирующий гормон Кожа (меланоциты) Пигментация Гормон паращитовидных желез Кости, почки Концентрация Са и Р в сыворотке крови Простациклин, простагландин Тромбоциты Агрегация тромбоцитов Тиреотропный гормон Щитовидная железа Продуцирование и высвобождение Тз и Т 4 Рилизинг-фактор тиреотропного гормона Гипофиз Высвоб-е тиреотропного гормона Вазопрессин Почки Концентрация мочи П 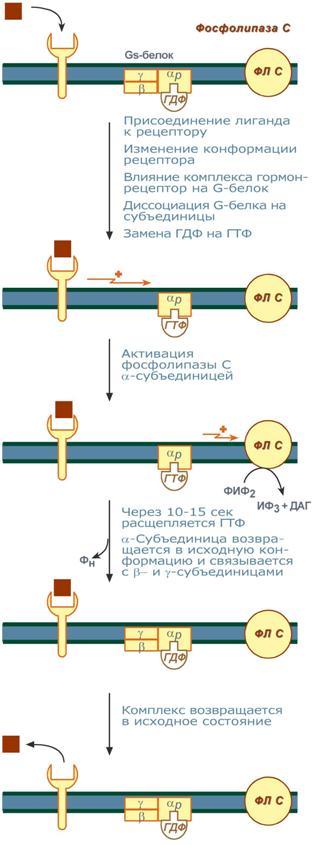 о этому механизму, который получил название кальций-фосфолипидный механизм, действуют вазопрессин (через V1-рецепторы), адреналин (через α1-адренорецепторы), ангиотензин II. о этому механизму, который получил название кальций-фосфолипидный механизм, действуют вазопрессин (через V1-рецепторы), адреналин (через α1-адренорецепторы), ангиотензин II.Принцип работы этого механизма совпадает с предыдущим, но вместо аденилатциклазы мишеневым ферментом для α-субъединицы служит фосфолипаза С. Фосфолипаза С расщепляет мембранный фосфолипид фосфатидилинозитолдифосфат (ФИФ2) до вторичных мессенджеров инозитолтрифосфата (ИФ3) и диацилглицерола (ДАГ). Этапы передачи сигнала: 1)Взаимодействие гормона с рецептором приводит к изменению конформации последнего. 2) Это изменение передается на G-белок, который состоит из трех субъединиц (αP, β и γ), α-субъединица связана с ГДФ. 3)В результате взаимодействия с рецептором β- и γ-субъединицы отщепляются, одновременно на αP-субъединице ГДФ заменяется на ГТФ. 4)Активированная таким образом αP-субъединица стимулирует фосфолипазу С, которая начинает расщепление ФИФ2 до двух вторичных мессенджеров – ИФ3 и ДАГ. 5)Инозитолтрифосфат открывает кальциевые каналы в эндоплазматическом ретикулуме, что вызывает увеличение концентрации ионов Cа2+. Диацилглицерол совместно с ионами Са2+ активирует протеинкиназу С. Кроме этого, диацилглицерол имеет и другую сигнальную функцию: он может распадаться на 1-моноацилглицерол и полиеновую жирную кислоту (обычно арахидоновую), из которой образуются эйкозаноиды. 6)Протеинкиназа С фосфорилирует ряд ферментов и в целом участвует в процессах клеточной пролиферации. Накопление ионов Са2+ в цитоплазме вызывает активацию определенных кальций-связывающих белков (например, кальмодулина). 7)Гидролиз ФИФ2 продолжается некоторое время, пока αP-субъединица, которая является ГТФ-азой, отщепляет фосфат от ГТФ. 8)Как только ГТФ превратился в ГДФ, то αP-субъединица инактивируется, теряет свое влияние на фосфолипазу C, обратно соединяется с β- и γ-субъединицами. Все возвращается в исходное положение. 9) Гормон отрывается от рецептора еще раньше: - если концентрация гормона в крови велика, то следующая его молекула присоединится к рецептору через малый промежуток времени и повторный запуск механизма произойдет быстро – в клетке активируются соответствующие процессы. 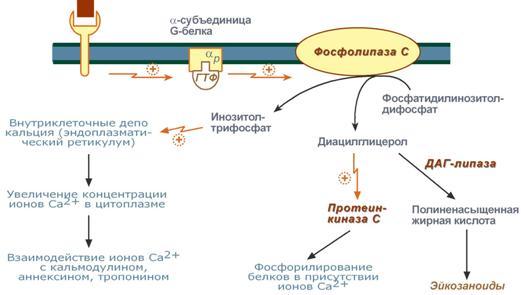 - если гормона в крови мало – для клетки наступает некоторая пауза, изменения метаболизма нет. - если гормона в крови мало – для клетки наступает некоторая пауза, изменения метаболизма нет. |