|
|
Ответы на билеты биохимия 2012. Белок это последовательность ак, связанных друг с другом пептидными связями
Патология: Гипофункция - Первичная недостаточность – болезнь Аддисона проявляется:
гипогликемия, повышенная чувствительность к инсулину, анорексия и снижение веса, слабость, гипотензия, гипонатриемия и гиперкалиемия, усиление пигментации кожи и слизистых (компенсаторное увеличение количества АКТГ, обладающего небольшим меланотропным действием).
Вторичная недостаточность возникает при дефиците АКТГ или снижении его эффекта на надпочечники – возникают все симптомы гипокортицизма, кроме пигментации.
Гиперфункция - Первичная – болезнь Кушинга проявляется:снижение толерантности к глюкозе – аномальная гипергликемия после сахарной нагрузки или после еды,гипергликемия из-за активации глюконеогенеза,ожирение лица и туловища (связано с повышенным влиянием инсулина при гипергликемии на жировую ткань) – буйволиный горбик, фартучный (лягушачий) живот, лунообразное лицо,глюкозурия,повышение катаболизма белков и повышение азота крови,остеопороз и усиление потерь кальция и фосфатов из костной ткани,снижение роста и деления клеток – лейкопения, иммунодефициты, истончение кожи, язвенная болезнь желудка и двенадцатиперстной кишки,нарушение синтеза коллагена и гликозаминогликанов,гипертония благодаря активации ренин-ангиотензиновой системы.Вторичная – синдром Иценко-Кушинга (избыток АКТГ) проявляется схоже с первичной формой.С интез минералокортикоидов в клетках клубочковой зоны коры надпочечников также начинается с превращения холестерола в прегненолон, а затем в прогестерон. Прогестерон гидроксилируется вначале по С21 с образованием 11-дезоксикортикостерона. Следующее гидроксилирование происходит по С11, что приводит к образованию кортикостерона, обладающего слабовыраженной глюкокортикоидной и минералокортикоидной активностью. В клетках клубочковой зоны 17-α-гидроксилаза отсутствует, но есть митохондриальная 18-гидроксилаза, при участии которой кортикостерон гидроксилируется, а затем дегидрируется с образованием альдегидной группы у С18. Основным гормоном у человека является альдостерон. Регуляция синтеза и секреции: Активируют:ангиотензин II, выделяемый при активации ренин-ангиотензиновой системы,повышение концентрации ионов калия в крови (связано с деполяризацией мембран, открытием кальциевых каналов и активацией аденилатциклазы). интез минералокортикоидов в клетках клубочковой зоны коры надпочечников также начинается с превращения холестерола в прегненолон, а затем в прогестерон. Прогестерон гидроксилируется вначале по С21 с образованием 11-дезоксикортикостерона. Следующее гидроксилирование происходит по С11, что приводит к образованию кортикостерона, обладающего слабовыраженной глюкокортикоидной и минералокортикоидной активностью. В клетках клубочковой зоны 17-α-гидроксилаза отсутствует, но есть митохондриальная 18-гидроксилаза, при участии которой кортикостерон гидроксилируется, а затем дегидрируется с образованием альдегидной группы у С18. Основным гормоном у человека является альдостерон. Регуляция синтеза и секреции: Активируют:ангиотензин II, выделяемый при активации ренин-ангиотензиновой системы,повышение концентрации ионов калия в крови (связано с деполяризацией мембран, открытием кальциевых каналов и активацией аденилатциклазы).
Активация ренин-ангиотензиновой системы. 1.Для активации этой системы существует два пусковых момента:
-снижение давления в приносящих артериолах почек, которое определяется барорецепторами клеток юкстагломерулярного аппарата. Причиной этого может быть любое нарушение почечного кровотока – атеросклероз почечных артерий, повышенная вязкость крови, обезвоживание, кровопотери и т.п.
-снижение концентрации ионов Na+ в первичной моче в дистальных канальцах почек, которое определяется осморецепторами клеток юкстагломерулярного аппарата. Возникает в результате бессолевой диеты, при длительном использовании диуретиков.
2. При выполнении одного или обоих пунктов клетки юкстагломерулярного аппарата активируются и из них в плазму крови секретируется фермент ренин.
3. Для ренина в плазме имеется субстрат – белок глобулиновой фракции ангиотензиноген.
4. В результате протеолиза от белка отщепляется декапептид под названием ангиотензин I.
5. Ангиотензин I при участии ангиотензин-превращающего фермента превращается в ангиотензин II.
6. Главными мишенями ангиотензина II служат кровеносные сосуды и клубочковая зона коры надпочечников.
7. Стимуляция кровеносных сосудов вызывает их спазм и восстановление артериального давления.
8. Из надпочечников после стимуляции секретируется альдостерон, действующий на дистальные канальцы почек.
9. При воздействии на канальцы почек увеличивается реабсорбция ионов Na+, вслед за натрием движется вода. В результате давление в системе вос-ся и концентрация ионов натрия увеличивается в плазме крови, а, значит и в первичной моче.
Транспорт кортикостеровдов. Кортизол в плазме крови находится в комплексе с α-глобулином транскортином и в небольшом количестве в свободной форме. Синтез транскортина протекает в печени и стимулируется эстрогенами. Т1/2 кортизола составляет 1,5-2 ч. Несвязанный, или свободный кортизол, составляет около 8% от общего количества гормона в плазме и является биологически активной фракцией. Альдостерон не имеет специфического транспортного белка, но образует слабые связи с альбумином. Катаболизм гормонов коры надпочечников происходит прежде всего в печени. Здесь протекают реакции гидроксилирования, окисления и восстановления гормонов. Продукты катаболизма кортикостеровдов (кроме кортикостерона и альдостерона) выводятся с мочой в форме 17-кетостероидов, образующихся в результате отщепления боковой цепи. Эти продукты метаболизма выделяются преимущественно в виде конъюгатов с глюкуроновой и серной кислотами. Определение 17-кетостероидов в моче позволяет оценить как количество глюкокортикоидов, секретируемых корой надпочечников, так и функцию надпочечников. Мишени и эффекты: Воздействует на слюнные железы, на дистальные канальцы и собирательные трубочки почек. В почках усиливает реабсорбцию ионов натрия и потерю ионов калия посредством следующих эффектов: увеличивает количество Na+,K+-АТФазы на базальной мембране эпителиальных клеток, стимулирует синтез митохондриальных белков и увеличение количества нарабатываемой в клетке энергии для работы Na+,K+-АТФазы, стимулирует образование Na-каналов на апикальной мембране клеток почечного эпителия.
Патология: Гипофункция -Не отмечена. Гиперфункция - Синдром Конна (первичный альдостеронизм) – возникает при аденомах клубочковой зоны. Характеризуется триадой признаков: гипертензия, гипернатриемия, алкалоз. Вторичный гиперальдостеронизм – гиперплазия и гиперфункция юкстагломерулярных клеток и избыточная секреция ренина и ангиотензина II.
53. Строение, синтез и метаболизм йодтиронинов. Влияние на обмен веществ. Гипо- и гипертиреозы: механизм возникновения и последствия.
В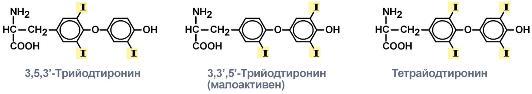 щитовидной железе синтезируются гормоны - йодированные производные тирозина. Они объединены общим названием йодтиронины. К ним относят 3,5,3'-трийодтиронин (трийодтиронин, Т3) и 3,5,3',5'-тетрайодтиронин (Т4), или тироксин. щитовидной железе синтезируются гормоны - йодированные производные тирозина. Они объединены общим названием йодтиронины. К ним относят 3,5,3'-трийодтиронин (трийодтиронин, Т3) и 3,5,3',5'-тетрайодтиронин (Т4), или тироксин.
Й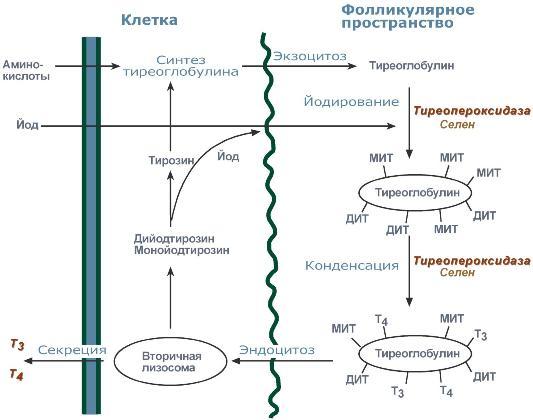 одтиронины синтезируются в составе белка тиреоглобулина (Тг) в фолликулах, которые представляют собой морфологическую и функциональную единицу щитовидной железы. Тиреоглобулин синтезируется в базальной части клеток, перемещается к просвету и хранится во внеклеточных коллоидах, при тиреотропной стимуляции пиноцитируется фолликулярными клетками, сливается с лизосомами и гидролизуется. Три- и тетрайодтиронин секретируются в кровь. В крови гормоны транспортируются специфическим глобулином, а также альбумином. Этапы синтеза тиреоидных гормонов: 1.Транспорт йода в клетки щитовидной железы: Йод в виде органических и неорганических соединений поступает в ЖКТ с пищей и питьевой водой. Суточная потребность в йоде составляет 150-200 мкг. 25-30% этого количества йодидов захватывается щитовидной железой. Транспорт йодида в кл щитовидной железы - энергозависимый процесс и происходит при участии транспортного белка против электрохимического градиента (соотношение концентраций I- в железе к концентрации I- в сыворотке крови в норме составляет 25:1). Работа этого йодид-переносящего белка сопряжена с Nа+,К+-АТФ-азой. Концентрационная способность железы регулируется ТТГ 2. Окисление йода. Окисление I- в I+ происходит при участии гемсодержащей тиреопероксидазы и Н2О2 в качестве окислителя. Окисление йодида до состояния с более высокой валентностью, что необходимо для органификации йодида и биосинтеза тиреоидных гормонов. 3. Йодирование тирозина (органификация йодида) – Окисленный йод взаимодействует с остатками тирозина в молекуле тиреоглобулина. Эта реакция также катализируется тиреопероксидазой. 4. Образование йодтиронинов. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием монойод-тирозинов (МИТ) и дийодтирозинов (ДИТ). Две молекулы ДИТ конденсируются с образованием йодтиронина Т4, а МИТ и ДИТ - с образованием йодтиронина Т3. Йодтиреоглобулин транспортируется из коллоида в фолликулярную клетку путём эндоцитоза и гидролизуется ферментами лизосом с освобождением Т3 и Т4. Если йода поступает достаточно, соотношение Т4/Т3=7:1. При недостатке, Т4/Т3 и ДИТ/МИТ снижается. Чем хуже железа обеспечена йодом, тем выше доля МИТ и Т3, что считается компенсаторным явлением. одтиронины синтезируются в составе белка тиреоглобулина (Тг) в фолликулах, которые представляют собой морфологическую и функциональную единицу щитовидной железы. Тиреоглобулин синтезируется в базальной части клеток, перемещается к просвету и хранится во внеклеточных коллоидах, при тиреотропной стимуляции пиноцитируется фолликулярными клетками, сливается с лизосомами и гидролизуется. Три- и тетрайодтиронин секретируются в кровь. В крови гормоны транспортируются специфическим глобулином, а также альбумином. Этапы синтеза тиреоидных гормонов: 1.Транспорт йода в клетки щитовидной железы: Йод в виде органических и неорганических соединений поступает в ЖКТ с пищей и питьевой водой. Суточная потребность в йоде составляет 150-200 мкг. 25-30% этого количества йодидов захватывается щитовидной железой. Транспорт йодида в кл щитовидной железы - энергозависимый процесс и происходит при участии транспортного белка против электрохимического градиента (соотношение концентраций I- в железе к концентрации I- в сыворотке крови в норме составляет 25:1). Работа этого йодид-переносящего белка сопряжена с Nа+,К+-АТФ-азой. Концентрационная способность железы регулируется ТТГ 2. Окисление йода. Окисление I- в I+ происходит при участии гемсодержащей тиреопероксидазы и Н2О2 в качестве окислителя. Окисление йодида до состояния с более высокой валентностью, что необходимо для органификации йодида и биосинтеза тиреоидных гормонов. 3. Йодирование тирозина (органификация йодида) – Окисленный йод взаимодействует с остатками тирозина в молекуле тиреоглобулина. Эта реакция также катализируется тиреопероксидазой. 4. Образование йодтиронинов. Под действием тиреопероксидазы окисленный йод реагирует с остатками тирозина с образованием монойод-тирозинов (МИТ) и дийодтирозинов (ДИТ). Две молекулы ДИТ конденсируются с образованием йодтиронина Т4, а МИТ и ДИТ - с образованием йодтиронина Т3. Йодтиреоглобулин транспортируется из коллоида в фолликулярную клетку путём эндоцитоза и гидролизуется ферментами лизосом с освобождением Т3 и Т4. Если йода поступает достаточно, соотношение Т4/Т3=7:1. При недостатке, Т4/Т3 и ДИТ/МИТ снижается. Чем хуже железа обеспечена йодом, тем выше доля МИТ и Т3, что считается компенсаторным явлением.
Регуляция синтеза и секреции: Активируют: тиреотропный гормон на этапах поглощения йода, синтеза тиреоглобулина, эндоцитоза и секреции Т3 и Т4 в кровь.Уменьшают: тироксин и трийодтиронин (по механизму обратной отрицательной связи), высокие концентрации йода в крови (бесконтрольный прием препаратов KJ).
Метаболизм йодтиронинов. От половины до двух третей Т3 и Т4 находятся в организме вне щитовидной железы. Большая часть их циркулирует в крови в связанной форме в комплексе с белками: тироксинсвязывающим глобулином (ТСГ) и тироксинсвязывающим преальбумином (ТСПА). ТСГ служит основным транспортным белком йодтиронинов, а также формой их депонирования. Он обладает более высоким сродством к Т3 и Т4 и в нормальных условиях связывает почти всё количество этих гормонов. Только 0,03% Т4 и 0,3% Т3 находятся в крови в свободной форме. Т1/2 Т4 в плазме в 4-5 раз больше, чем Т3. Для Т4 этот период составляет около 7 дней, а для Т3 - 1-1,5 дня. Биологическая активность йодтиронинов обусловлена несвязанной фракцией. Т3 - основная биологически активная форма йодтиронинов; его сродство к рецептору клеток-мишеней в 10 раз выше, чем у Т4. Другие пути метаболизма йодтиронинов включают полное дейодирование, дезаминирование или декарбоксилирование. Йодированные продукты катаболизма йодтиронинов конъюгируют-ся в печени с глюкуроновой или серной кислотами, секретируются с жёлчью, в кишечнике вновь всасываются, дейодируются в почках и выделяются с мочой. Механизм действия: Цитозольный.
Мишени и эффекты: Клетки-мишени йодтиронинов имеют 2 типа рецепторов к этим гормонам. Основные эффекты йодтиронинов - результат их взаимодействия с высокоспецифичными рецепторами, которые в комплексе с гормонами постоянно находятся в ядре и взаимодействуют с определёнными последовательностями ДНК, участвуя в регуляции экспрессии генов. Другие рецепторы расположены в плазматической мембране клеток, но это не те же самые белки, что в ядре. Они обладают более низким сродством к йодтиронинам и обеспечивают связывание гормонов для удержания их в непосредственной близости к клетке. Рецепторы к йодтиронинам имеют все ткани организма. В клетках-мишенях тироксин дейодируется и активной формой является трийодтиронин (3,5,3'-производное). Эта реакция ослаблена у плода, новорожденных и престарелых. Главным эффектом трийодтиронина является повышение активности Na+,K+-АТФазы, что приводит к быстрому расходованию АТФ и по механизму дыхательного контроля запускает катаболизм углеводов и липидов. В митохондриях увеличивается количество АТФ/АДФ-транслоказы и потребление кислорода. Сопутствующим эффектом усиления катаболизма является наработка тепла.
Белковый обмен: Усиливает транспорт аминокислот в клетки. Активирует синтез дифференцировочных белков в ЦНС, гонадах, костной ткани и обусловливает развитие этих тканей.
Углеводный обмен: Увеличивает гликогенолиз и аэробное окисление глюкозы.
Липидный обмен: Стимулирует липолиз, β-окисление жирных кислот, подавляет стероидогенез.
Нуклеиновый обмен: Активирует начальные стадии синтеза пуринов и синтеза пиримидинов, стимулирует дифференцировочный синтез РНК и ДНК.
Также трийодтиронин усиливает выделение соматолиберина, что стимулирует секрецию гормона роста, также опосредует его метаболические эффекты. Этот эффект обеспечивает у детей в целом анаболическое действие тиреоидных гормонов. У взрослых действие тиреоидных гормонов в целом катаболическое, в надпочечниках подавляет синтез катехоламинов.
Патология. Гипофункция. Причина. Развивается при снижении синтеза гормонов в результате недостаточной стимуляции со стороны гипофиза и гипоталамуса, при заболевании самой железы, при нехватке необходимых веществ (аминокислоты, йод, селен). В 90% случаев причиной выраженного гипотиреоза является болезнь Хашимото, при которой вырабатываются блокирующие антиантитела к рецепторам ТТГ. Клиническая картина. Симптомами субклинического гипотиреоза, зачастую неспецифическими, могут быть отечность, сухость кожи и волос, брадикардия, легкое увеличение массы тела, повышение диастолического давления (точнее – понижение систолического), вялость, сонливость, запоры, чувствительность к холоду, снижение утренней температуры тела до 36,0°-35,5°С и ниже, бледность, скованность мышц, психическая инертность, апатия. У подростков отмечается отставание в физическом развитии, позднее половое созревание, функциональная дебильность, т.е. замедление мышления, снижение успеваемости в школе, неспособность к творческой деятельности, утрата чувства юмора. При наличии выраженного гипотиреоза у плодов, новорожденных и детей младшего возраста развивается кретинизм. При возникновении выраженного гипотиреоза у взрослых отмечается микседема, у женщин – бесплодие и галакторея, у обоих полов – деменция, психоз.
Гиперфункция. Причина. 90% случаев выраженного гипертиреоза вызвано наличием активирующих антиантител к рецепторам ТТГ. В этом случае заболевание носит название болезнь фон Базедова (в отечественной и европейской литературе), болезнь Грейвса (в американской литературе).
Клиническая картина. Симптомами гипертиреоза являются субфебрильная температура (до 37,5°С, особенно к вечеру), нервное возбуждение, эмоциональная лабильность и нервозность (плаксивость), похудание, мышечная слабость, повышение аппетита, тахикардия, потливость, непереносимость жары, тремор, диарея.
54. Катехоламины. Синтез, депонирование и метаболизм катехоламинов. Механизм действия. Влияние на обмен веществ.
Подобно задней доле гипофиза, мозговой слой надпочечников - производное нервной ткани. Его можно рассматривать как продолжение симпатической нервной системы, так как преганглионарные волокна чревного нерва оканчиваются на хромаффинных клетках мозгового слоя надпочечников. При стимуляции преганглионарного нейрона хромаффинные клетки продуцируют катехоламины (они относятся к группе фенилалкиламинов) - адреналин (80%), норадреналин (20%), дофамин (менее1%). По химическому строению катехоламины - 3,4-дигидроксипроизводные фенилэтиламина. Непосредственным предшественником гормонов служит тирозин. Собственно гормоном можно назвать только адреналин, поскольку два других катехоламина играют в основном медиаторную роль: норадреналин – в симпатической нервной системе, дофамин – в центральной. Адреналин относят к медиаторам симпатической и центральной нервной системы, а также к стресс-гормонам.
С интез катехоламинов происходит в цитоплазме и гранулах клеток мозгового слоя надпочечников. В гранулах происходит также запасание катехоламинов. Катехоламины поступают в гранулы путём АТФ-зависимого транспорта и хранятся в них в комплексе с АТФ в соотношении 4:1 (гормон-АТФ). Разные гранулы содержат разные катехоламины: некоторые только адреналин, другие - норадреналин, третьи - оба гормона. Синтез осуществляется в клетках мозгового слоя надпочечников (80% всего адреналина), синтез норадреналина (80%) происходит также в нервных синапсах. Секреция гормонов из гранул происходит путём экзоцитоза. Катехоламины и АТФ освобождаются из гранул в том же соотношении, в каком они сохраняются в гранулах. В отличие от симпатических нервов, клетки мозгового слоя надпочечников лишены механизма обратного захвата выделившихся катехоламинов. интез катехоламинов происходит в цитоплазме и гранулах клеток мозгового слоя надпочечников. В гранулах происходит также запасание катехоламинов. Катехоламины поступают в гранулы путём АТФ-зависимого транспорта и хранятся в них в комплексе с АТФ в соотношении 4:1 (гормон-АТФ). Разные гранулы содержат разные катехоламины: некоторые только адреналин, другие - норадреналин, третьи - оба гормона. Синтез осуществляется в клетках мозгового слоя надпочечников (80% всего адреналина), синтез норадреналина (80%) происходит также в нервных синапсах. Секреция гормонов из гранул происходит путём экзоцитоза. Катехоламины и АТФ освобождаются из гранул в том же соотношении, в каком они сохраняются в гранулах. В отличие от симпатических нервов, клетки мозгового слоя надпочечников лишены механизма обратного захвата выделившихся катехоламинов.
В плазме крови катехоламины образуют непрочный комплекс с альбумином. Адреналин транспортируется в основном к печени и скелетным мышцам. Норадреналин образуется в основном в органах, иннервируемых симпатическими нервами (80% от общего количества). Норадреналин лишь в незначительных количествах достигает периферических тканей. Основная часть катехоламинов быстро метаболизируется в различных тканях при участии специфических. Лишь небольшая часть адреналина (
5%) выделяется с мочой. Регуляция синтеза и секреции: Активируют: стимуляция чревного нерва, стресс. Уменьшают: гормоны щитовидной железы. Катехоламины действуют на клетки-мишени через рецепторы, локализованные в плазматической мембране. Механизм действия гормонов разный в зависимости от рецептора (R). Конечный эффект гормонов зависит от преобладания типа рецепторов на клетке и концентрации гормона в крови. Выделяют 2 главных класса таких рецепторов: α-адренергические и β-адренергические. Все рецепторы катехоламинов - гликопротеины, передают сигналы в клетки с помощью разных вторичных посредников. Это определяет характер их влияния на метаболизм клеток-мишеней. Адреналин взаимодействует как с α-R, так и с β-R; норадреналин в физиологических концентрациях главным образом взаимодействует с α-R. Аденилатциклазный механизм: а) при задействовании α2-адренорецепторов аденилатциклаза ингиб-ся,
б) при задействовании β1- и β2-адренорецепторов аденилатциклаза активируется. Кальций-фосфолипидный механизм: при возбуждении α1-адренорецепторов.Мишени и эффекты: 1)При возбуждении α1-адренорецепторов (печень, СС и мочеполовая системы): активация гликогенолиза, сокращение гладких мышц кровеносных сосудов и мочеполовой системы.2) При возбуждении α2-адренорецепторов (жировая ткань, поджелудка, почки):подавление липолиза (уменьшение стимуляции ТАГ-липазы), подавление секреции инсулина и секреции ренина. 3) При возбуждении β1-адренорецепторов (есть во всех тк): активация липолиза, увеличение силы и ЧСС миокарда. 4)При возбуждении β2-адренорецепторов (есть во всех тк): стимуляция гликогенолиза в печени и мышцах, и глюконеогенеза в печени, расслабление гладких мышц бронхов, кровеносных сосудов, мочеполовой системы и ЖКТ. В целом катехоламины отвечают за биохимические реакции адаптации к острым стрессам, связанным с мышечной активностью – "борьба или бегство": -усиление липолиза и продукция жирных кислот в жировой ткани для мышечной активности, -гипергликемия за счет глюконеогенеза и гликогенолиза в печени для повышения устойчивости ЦНС, -стимуляция гликогенолиза в мышцах, -активация протеолиза в лимфоидной ткани для обеспечения глюконеогенеза субстратом (АК), -снижение анаболических процессов через уменьшение секреции инсулина.
Адаптация также прослеживается в физиологических реакциях: -мозг – усиление кровотока и стимуляция обмена глюкозы,
-мышцы – усиление сократимости, -ССС – увеличение силы и ЧСС миокарда, -легкие – расширение бронхов, улучшение вентиляции и потребления О2, -кожа – снижение кровотока. Удаление катехоламинов из синаптич. щели происходит путем обратного захвата в пресинаптич. окончания. Этот активный транспорт происходит с использованием энергии градиента Na+ на мембране. Часть катехоламинов диффундирует из синапса в межклеточное пространство и затем поступает в кровь. Инактивируются катехоламины путем окислит, дезаминирования, катализируемого катехол-О-метилтрансферазой. Метаболиты катехоламинов удаляются из организма почками, поэтому их содержание в моче может указывать на активность симпатич. НС и мозгового слоя надпочечников.
Основная патология мозгового вещества надпочечников - феохромоцитома, опухоль, образованная хромаффинными клетками и продуцирующая катехоламины. Клинически феохромоцитома проявляется повторяющимися приступами головной боли, сердцебиения, потливости, повышением АД и сопровождается характерными изменениями метаболизма.
|
|
|
 Скачать 5.15 Mb.
Скачать 5.15 Mb.