Ответы на билеты биохимия 2012. Белок это последовательность ак, связанных друг с другом пептидными связями
 Скачать 5.15 Mb. Скачать 5.15 Mb.
|

|
57. Регуляция обмена кальция и фосфора. Роль паратгормона и тиреокальцитонина. Витамин Д. Роль 1,25-дигидроксикальциферола в регуляции кальция и фосфатов. Рахит. Концентрация кальция во внеклеточной жидкости в норме поддерживается на строго постоянном уровне, редко повышаясь или снижаясь на несколько процентов относительно нормальных величин, составляющих 9,4 мг/дл, что эквивалентно 2,4 ммоль кальция на литр. Такой строгий контроль очень важен в связи с основной ролью кальция во многих физиологических процессах, включая сокращение скелетных, сердечной и гладких мышц, свертывание крови, передачу нервных импульсов. Возбудимые ткани, в том числе нервная, очень чувствительны к изменениям концентрации кальция, и увеличение концентрации ионов кальция по сравнению с нормой (гипсркальциемия) вызывает нарастающее поражение нервной системы; напротив, снижение концентрации кальция (гипокальциемия) повышает возбудимость нервной системы. Важная особенность регуляции концентрации внеклеточного кальция: только около 0,1% общего количества кальция организма присутствует во внеклеточной жидкости, около 1 % — находится внутри клеток, а остальное количество хранится в костях, поэтому кости могут рассматриваться в качестве большого хранилища кальция, выделяющего его во внеклеточное пространство, если концентрация кальция там снижается, и, напротив, забирающего избыток кальция на хранение. Приблизительно 85% фосфатов организма хранится в костях, от 14 до 15% — в клетках, и только менее 1% присутствует во внеклеточной жидкости. Концентрация фосфатов во внеклеточной жидкости не так строго регулируется, как концентрация кальция, хотя они выполняют разнообразные важные функции, контролируя многие процессы совместно с кальцием. Всасывание кальция и фосфатов в кишечнике и их экскреция с калом. Обычная скорость поступления кальция и фосфатов составляет приблизительно 1000 мг/сут, что соответствует количеству, извлекаемому из 1 л молока. Обычно двухвалентные катионы, такие как ионизированный кальций, плохо абсорбируются в кишечнике. Однако, как обсуждается далее, витамин D способствует всасыванию кальция в кишечнике, и почти 35% (около 350 мг/сут) потребленного кальция абсорбируется. Оставшийся в кишечнике кальций попадает в каловые массы и удаляется из организма. Дополнительно около 250 мг/сут кальция попадает в кишечник в составе пищеварительных соков и слущивающихся клеток. Таким образом, около 90% (900 мг/сут) из ежесуточного поступления кальция выводится с калом. Гипокальциемия вызывает возбуждение нервной системы и тетанию. Если концентрация ионов кальция во внеклеточной жидкости падает ниже нормальных значений, нервная система постепенно становится все более возбудимой, т.к. это изменение приводит к повышению проницаемости для ионов натрия, облегчая генерацию потенциала действия. В случае падения концентрации ионов кальция до уровня, составляющего 50% нормы, возбудимость периферических нервных волокон становится так велика, что они начинают спонтанно разряжаться. Гиперкальциемия понижает возбудимость нервной системы и мышечную активность. Если концентрация кальция в жидких средах организма превышает норму, возбудимость нервной системы снижается, что сопровождается замедлением рефлекторных ответов. Увеличение концентрации кальция приводит к снижению интервала QT на электрокардиограмме, снижению аппетита и запорам, возможно, вследствие снижения контрактильной активности мышечной стенки гастроинтестинального тракта. Эти депрессивные эффекты начинают проявляться, когда уровень кальция поднимается выше 12 мг/дл, и становятся заметными, когда уровень кальция превышает 15 мг/дл. Формирующиеся нервные импульсы достигают скелетных мышц, вызывая тетанические сокращения. Следовательно, гипокальциемия вызывает тетанию, иногда она провоцирует эпилептиформные приступы, поскольку гипокальциемия повышает возбудимость мозга. Всасывание фосфатов в кишечнике осуществляется легко. Кроме тех количеств фосфатов, которые выводятся с калом в виде солей кальция, почти все содержащиеся в дневном рационе фосфаты всасываются из кишечника в кровь и затем экскретируются с мочой. Экскреция кальция и фосфатов почкой. Приблизительно 10% (100 мг/сут) поступившего в организм кальция экскретируются с мочой, около 41% кальция в плазме связано с белками и поэтому не фильтруется из гломерулярных капилляров. Оставшееся количество объединяется с анионами, например с фосфатами (9%), или ионизируется (50%) и фильтруется клубочками в почечные канальцы. В норме в канальцах почки реабсорбируется 99% отфильтрованного кальция, поэтому в сутки экскретируются с мочой почти 100 мг кальция. Приблизительно 90% кальция, содержащегося в гломерулярном фильтрате, реабсорбируется в проксимальных канальцах, петле Генле и в начале дистальных канальцев. Затем в конце дистальных канальцев и в начале собирательных протоков реабсорбируются оставшиеся 10% кальция. Реабсорбция становится высокоизбирательной и зависит от концентрации кальция в крови. Если концентрация кальция в крови низка, реабсорбция возрастает, в итоге кальций почти не теряется с мочой. Напротив, когда концентрация кальция в крови незначительно превышает нормальные значения, экскреция кальция значительно увеличивается. Наиболее важным фактором, контролирующим реабсорбцию кальция в дистальных отделах нефрона и, следовательно, регулирующим уровень экскреции кальция, является паратгормон. Почечная экскреция фосфатов регулируется механизмом обильного потока. Это означает, что когда концентрация фосфатов в плазме снижается ниже критического значения (около 1 ммоль/л), все фосфаты из гломеруляр-ного фильтрата реабсорбируются и перестают выводиться с мочой. Но если концентрация фосфатов превышает значение нормы, потери его с мочой прямо пропорциональны дополнительному увеличению его концентрации. Почки регулируют концентрацию фосфатов в экстрацеллюлярном пространстве, изменяя скорость экскреции фосфатов соответственно их концентрации в плазме и скорости фильтрации фосфатов в почке. Однако, как мы увидим далее, паратгормон может существенно увеличить экскрецию фосфатов почками, поэтому он играет важную роль в регуляции концентрации фосфатов в плазме наряду с контролем концентрации кальция. Паратгормон является мощным регулятором концентрации кальция и фосфатов, осуществляющим свои влияния, управляя процессами реабсорбции в кишечнике, экскрецией в почке и обменом этих ионов между внеклеточной жидкостью и костью. Избыточная активность паращитовидных желез вызывает быстрое вымывание солей кальция из костей с последующим развитием гиперкальциемии во внеклеточной жидкости; напротив, гипофункция паращитовидных желез приводит к гипокальциемиии, часто — с развитием тетании. Функциональная анатомия паращитовидных желез. В норме у человека существуют четыре паращитовидные железы. Они расположены сразу после щитовидной железы, попарно у верхнего и нижнего ее полюсов. Каждая паращитовидная железа является образованием около 6 мм длиной, 3 мм шириной и 2 мм высотой. Макроскопически паращитовидные железы выглядят как темный бурый жир, определить их местонахождение во время операции на щитовидной железе затруднительно, т.к. они часто выглядят, как дополнительная доля щитовидной железы. Именно поэтому до момента, когда была установлена важность этих желез, тотальная или субтотальная тиреоидэктомия заканчивалась одновременным удалением паращитовидных желез. Удаление половины околощитовидных желез не вызывает серьезных физиологических нарушений, удаление трех или всех четырех желез приводит к транзиторному гипопаратиреоидизму. Но даже небольшое количество оставшейся ткани паращитовидной железы способно за счет гиперплазии обеспечить нормальную функцию паращитовидных желез. Паратиреоидные железы взрослого человека состоят преимущественно из главных клеток и из большего или меньшего количества оксифильных клеток, которые отсутствуют у многих животных и у молодых людей. Главные клетки предположительно секретируют большее, если не все количество паратгормона, а у оксифильных клеток — свое предназначение. Считается, что они являются модификацией или исчерпавшей свой ресурс формой главных клеток, которые больше не синтезируют гормон. Химическая структура паратгормона. ПТГ выделен в очищенном виде. Первоначально он синтезируется на рибосомах в виде препрогормона, полипептидной цепочки из ПО аминокислотных остатков. Затем расщепляется до прогормона, состоящего из 90 аминокислотных остатков, потом — до стадии гормона, который включает 84 аминокислотных остатка. Процесс этот осуществляется в эндоплазматическом ретикулуме и аппарате Гольджи. В итоге гормон упаковывается в секреторные гранулы в цитоплазме клеток. Окончательная форма гормона имеет молекулярную массу 9500; более мелкие соединения, состоящие из 34 аминокислотных остатков, примыкающие к N-концу молекулы паратгормона, также выделенные из паращитовидных желез, обладают активностью ПТГ в полной мере. Установлено, что почки полностью выводят форму гормона, состоящую из 84 аминокислотных остатков, очень быстро, в течение нескольких минут, в то время как оставшиеся многочисленные фрагменты длительное время обеспечивают поддержание высокой степени гормональной активности. Тиреокальцитонин — гормон, вырабатываемый у млекопитающих и у человека парафолликулярными клетками щитовидной железой, паращитовидной железой и вилочковой железой. У многих животных, например, рыб, аналогичный по функциям гормон производится не в щитовидной железе (хотя она есть у всех позвоночных животных), а в ултимобранхиальных тельцах и потому называется просто кальцитонином. Тиреокальцитонин принимает участие в регуляции фосфорно-кальциевого обмена в организме, а также баланса активности остеокластов и остеобластов, функциональный антагонист паратгормона. Тиреокальцитонин понижает содержание кальция и фосфата в плазме крови за счёт усиления захвата кальция и фосфата остеобластами. Он также стимулирует размножение и функциональную активность остеобластов. Одновременно тиреокальцитонин тормозит размножение и функциональную активность остеокластов и процессы резорбции кости. Тиреокальцитонин является белково-пептидным гормоном, с молекулярной массой3600. Усиливает отложение фосфорно-кальциевых солей на коллагеновую матрицу костей. Тиреокальцитонин, как и паратгормон, усиливает фосфатурию. Кальцитриол Строение: Представляет собой производное витамина D и относится к стероидам.  Синтез: Образующийся в коже под действием ультрафиолета и поступающие с пищей холекальциферол (витамин D3) и эргокальциферол (витамин D2) гидроксилируются в печени по С25 и в почках по С1. В результате формируется 1,25-диоксикальциферол (кальцитриол). Регуляция синтеза и секреции Активируют: Гипокальциемия повышает гидроксилирование по С1 в почках. Уменьшают: Избыток кальцитриола подавляет гидроксилирование по С1 в почках. Механизм действия: Цитозольный. Мишени и эффекты: Эффект кальцитриола заключается в увеличении концентрации кальция и фосфора в крови: в кишечнике индуцирует синтез белков, отвечающих за всасывание кальция и фосфатов, в почках повышает реабсорбцию кальция и фосфатов, в костной ткани усиливает резорбцию кальция. Патология: Гипофункция Соответствует картине гиповитаминоза D. Роль 1.25-дигидроксикальци-ферола в обмене Ca и P.: Усиливает всасывание Ca и P из кишечника, Усиливает реабсорбцию Ca и P почками, Усиливает минерализацию молодой кости, Стимулирует остеокласты и выход Ca из старой кости. Витамин D (кальциферол, антирахитический) Источники: Имеется два источника поступления витамина D: печень, дрожжи, жирномолочные продукты (сливочное масло, сливки, сметана), желток яиц, образуется в коже при ультрафиолетовом облучении из 7-дегидрохолестерола в количестве 0,5-1,0 мкг/сут. Суточная потребность: Для детей – 12-25 мкг или 500-1000 МЕ, у взрослых потребность гораздо меньше. С 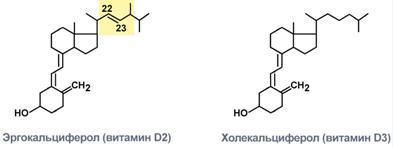 троение: Витамин представлен двумя формами – эргокальциферол и холекальциферол. Химически эргокальциферол отличается от холекальциферола наличием в молекуле двойной связи между С22 и С23 и метильной группой при С24. троение: Витамин представлен двумя формами – эргокальциферол и холекальциферол. Химически эргокальциферол отличается от холекальциферола наличием в молекуле двойной связи между С22 и С23 и метильной группой при С24.После всасывания в кишечнике или после синтеза в коже витамин попадает в печень. Здесь он гидроксилируется по С25 и кальциферолтранспортным белком переносится к почкам, где еще раз гидроксилируется, уже по С1. Образуется 1,25-дигидроксихолекальциферол или кальцитриол. Реакция гидроксилирования в почках стимулируется паратгормоном, пролактином, соматотропным гормоном и подавляется высокими концентрациями фосфатов и кальция. Биохимические функции: 1. Увеличение концентрации кальция и фосфатов в плазме крови.Для этого кальцитриол:стимулирует всасывание ионов Ca2+ и фосфат-ионов в тонком кишечнике (главная функция),стимулирует реабсорбцию ионов Ca2+ и фосфат-ионов в проксимальных почечных канальцах. 2. В костной ткани роль витамина D двояка: стимулирует выход ионов Ca2+ из костной ткани, так как способствует дифференцировке моноцитов и макрофагов в остеокласты и снижению синтеза коллагена I типа остеобластами, повышает минерализацию костного матрикса, так как увеличивает производство лимонной кислоты, образующей здесь нерастворимые соли с кальцием. 3. Участие в реакциях иммунитета, в частности в стимуляции легочных макрофагов и в выработке ими азотсодержащих свободных радикалов, губительных, в том числе, для микобактерий туберкулеза. 4. Подавляет секрецию паратиреоидного гормона через повышение концентрации кальция в крови, но усиливает его эффект на реабсорбцию кальция в почках. Гиповитаминоз. Приобретенный гиповитаминоз.Причина. Часто встречается при пищевой недостаточности у детей, при недостаточной инсоляции у людей, не выходящих на улицу или при национальных особенностях одежды. Также причиной гиповитаминоза может быть снижение гидроксилирования кальциферола (заболевания печени и почек) и нарушение всасывания и переваривания липидов (целиакия, холестаз). Клиническая картина: У детей от 2 до 24 месяцев проявляется в виде рахита, при котором, несмотря на поступление с пищей, кальций не усваивается в кишечнике, а в почках теряется. Это ведет к снижению концентрации кальция в плазме крови, нарушению минерализации костной ткани и, как следствие, к остеомаляции (размягчение кости). Остеомаляция проявляется деформацией костей черепа (бугристость головы), грудной клетки (куриная грудь), искривление голени, рахитические четки на ребрах, увеличение живота из за гипотонии мышц, замедляется прорезывание зубов и зарастание родничков. У взрослых тоже наблюдается остеомаляция, т.е. остеоид продолжает синтезироваться, но не минерализуется. Развитие остеопороза частично также связывают с витамин D- недостаточностью. Наследственный гиповитаминоз Витамин D-зависимый наследственный рахит I типа, при котором имеется рецессивный дефект почечной α1-гидроксилазы. Проявляется задержкой развития, рахитическими особенностями скелета и т.д. Лечение – препараты кальцитриола или большие дозы витамина D. Витамин D-зависимый наследственный рахит II типа, при котором наблюдается дефект тканевых рецепторов кальцитриола. Клинически заболевание схоже с I типом, но дополнительно отмечаются аллопеция, milia, эпидермальные кисты, мышечная слабость. Лечение варьирует в зависимости от тяжести заболевания, помогают большие дозы кальциферола. Гипервитаминоз. Причина Избыточное потребление с препаратами (не менее 1,5 млн МЕ в сутки). Клиническая картина: Ранними признаками передозировки витамина D являются тошнота, головная боль, потеря аппетита и веса тела, полиурия, жажда и полидипсия. Могут быть запоры, гипертензия, мышечная ригидность.Хронический избыток витамина D приводит к гипервитаминозу, при котором отмечается:деминерализация костей, приводящая к их хрупкости и переломам.увеличение концентрации ионов кальция и фосфора в крови, приводящее к кальцификации сосудов, ткани легких и почек. Лекарственные формы Витамин D – рыбий жир, эргокальциферол, холекальциферол. 1,25-Диоксикальциферол (активная форма) – остеотриол, оксидевит, рокальтрол, форкал плюс. 58. Гормоны, производные жирных кислот. Синтез. Функции. По химической природе гормональные молекулы относят к трем группам соединений: 1)белки и пептиды; 2) производные аминокислот; 3) стероиды и производные жирных кислот. К эйкозаноидам (είκοσι, греч.-двадцать) относят окисленные производные эйкозановых к-т: эйкозотриеновой (С20:3), арахидоновой (С20:4), тимнодоновой (С20:5) ж-х к-т. Активность эйкозаноидов значительно разнится от числа двойных связей в молекуле, которое зависит от строения исходной ж-ой к-ы. Эйкозаноиды называют гормоноподобными вещ-ми, т.к. они могут оказывать только местное действие, сохраняясь в крови в течение неск-х сек. Обр-ся во всех органах и тканях практически всеми типами кл. Депонироваться эйкозаноиды не могут, разрушаются в течение неск-их сек, и поэтому кл должна синтезировать их постоянно из поступающих жирных кислот ω6- и ω3-ряда. Выделяют три основные группы: Простагландины (Pg) – синтезируются практически во всех клетках, кроме эритроцитов и лимфоцитов. Выделяют типы простагландинов A, B, C, D, E, F. Функции простагландинов сводятся к изменению тонуса гладких мышц бронхов, мочеполовой и сосудистой системы, желудочно-кишечного тракта, при этом направленность изменений различна в зависимости от типа простагландинов, типа клетки и условий. Они также влияют на температуру тела. Могут активировать аденилатциклазу Простациклины являются подвидом простагландинов (Pg I), вызывают дилатацию мелких сосудов, но еще обладают особой функцией – ингибируют агрегацию тромбоцитов. Их активность возрастает при увеличении числа двойных связей. Синтезируются в эндотелии сосудов миокарда, матки, слизистой желудка. Тромбоксаны (Tx) образуются в тромбоцитах, стимулируют их агрегацию и вызывают сужение сосудов. Их активность снижается при увеличении числа двойных связей. Увеличивают активность фосфоинозитидного обмена Лейкотриены (Lt) синтезируются в лейкоцитах, в клетках легких, селезенки, мозга, сердца. Выделяют 6 типов лейкотриенов A, B, C, D, E, F. В лейкоцитах они стимулируют подвижность, хемотаксис и миграцию клеток в очаг воспаления, в целом они активируют реакции воспаления, предотвращая его хронизацию. Также вызывают сокращение мускулатуры бронхов (в дозах в 100-1000 раз меньших, чем гистамин). повышают проницаемость мембран для ионов Са2+. Поскольку цАМФ и ионы Са2+ стимулируют синтез эйкозаноидов, замыкается положительная обратная связь в синтезе этих специфических регуляторов. |