Ответы на билеты биохимия 2012. Белок это последовательность ак, связанных друг с другом пептидными связями
 Скачать 5.15 Mb. Скачать 5.15 Mb.
|

|
Процессинг предшественника транспортной РНК. 1. Модификация нуклеотидов в молекуле путем дезаминирования, метилирования, восстановления. Например, образование псевдоуридина и дигидроуридина. 2. Формирование антикодоновой петли происходит путем сплайсинга и удаления интрона в средней части пре-тРНК. 3. Формирование на 3'-конце последовательности ЦЦА. Для этого у одних пре-тРНК с 3'-конца удаляются лишние нуклеотиды до "обнажения" триплета ЦЦА, у других идет присоединение этой последовательности. Рибосома-органелла клетки, участвующая в биосинтезе белка, состоит из молекул РНК (т. наз. рибосомные рибонуклеиновые кислоты, или рРНК) и белков. Осн. масса Р. локализована в цитоплазме. В бактериальной клетке Р. составляют до 30% ее сухой массы: на одну бактериальную клетку приходится примерно 104 Р. Состоят из двух разделяемых субчастиц, или рибосомных субъединиц. При определенных условиях (напр., при понижении концентрации Mg2 + в среде) Р. обратимо диссоциирует на две субчастицы с соотношением их мол. масс ок. 2:1. Прокарйотическая 70S Р. диссоциирует на субъединицы с коэф. седиментации 50S (мол. м. 1,5•106) и 30S (мол. м. 0,85•106). Эукариотическая Р. разделяется на субчастицы 60S и 40S. Две рибосомные субчастицы объединены в полную Р. строго определенным образом, предполагающим специфич. контакты их поверхностей. Полисома - Временный комплекс (4-5 и более) рибосом, транслирующих одновременно одну молекулу мРНК. Наличие в цитоплазме клеток значительного количества полисом свидетельствует о высокой интенсивности синтеза белка в конкретный момент времени. Много рибосом одновременно участв в синтезе белка на одной и той же мРНК, образ комплекс-полисому. Полирибосомы могут сущ в виде частиц, плавающих в цитоплазме клеток. Полирибосомы. По причине того, что продолжительность жизни матричной РНК невелика, перед клеткой стоит задача использовать ее максимально эффективно, т.е. получить максимальное количество "белковых копий". Для достижения этой цели на каждой мРНК может располагаться не одна, а несколько рибосом, встающих последовательно друг за другом и синтезирующих пептидные цепи. Такие образования называются полирибосомы. А где аминоацил-тРНК-синтетаза - фермент, АК - активируемая аминокислота, АТФ - аденозинтрифосфат, ПФ - пирофосфат, АМФ - аденозинмонофосфат. Карбоксильная группа аминокислоты в АК-АМФ связана ангидридной связью с остатком фосфорной к-ты (см. ф-лу; R'-ацил аминокислоты, R = Н), а в АК - тРНК - сложноэфирной связью с гидроксильной группой в положении 2' или 3' рибозы концевого остатка аденозина тРНК . Каждая аминоацил-тРНК-синтетаза специфична только по отношению к одной из 20 аминокислот, входящих в белки, и к одной или нескольким тРНК. Для тех аминокислот, для которых открыты две и более тРНК, соответствующая аминоацил-тРНК-синтетаза катализирует аминоацилирование всех этих тРНК. Это обстоятельство чрезвычайно важно, поскольку в дальнейшем в белковом синтезе «узнавание» аминоацил-тРНК основано не на природе аминокислоты, а на химической природе антикодона тРНК. Считается, что в молекуле каждой аминоацил-тРНК-синтетазы имеется по крайней мере 3 центра связывания: для аминокислоты, тРНК и АТФ. Аминоацил-тРНК-синтетазы в активном центре содержат гистидин, имидазольное кольцо которого участвует в связывании АТФ посредством ионов Mg2+. Специфичность р-ций, катализируемых аминоацил-тРНК-синтетазами, очень высока, что определяет точность белкового синтеза в живой клетке. Если аминоацил-тРНК-синтетаза осуществит ошибочное аминоацилирование тРНК близкой по структуре аминокислотой, произойдет коррекция путем катализируемого той же аминоацил-тРНК-синтетазой гидролиза ошибочных АК-тРНК до АК и тРНК. В цитоплазме содержится полный набор аминоацил-тРНК-синтетаз, кодируемых ядерным геномом; в хлоропластах и митохондриях есть свои аминоацил-тРНК-синтетазы. 40. Биологический код. Основные компоненты белоксинтезирующей системы. Биосинтез белка. Механизм. Адапторная функция тРНК и роль мРНК в этом процессе. Генетический (биологический) код – это способ кодирования информации о строении белков в виде нуклеотидной последовательности. Он предназначен для перевода четырехзначного языка нуклеотидов (А, Г, У, Ц) в двадцатизначный язык аминокислот. Он обладает характерными особенностями: Триплетность – три нуклеотида формируют кодон, кодирующий аминокислоту. Всего насчитывают 61 смысловой кодон. Специфичность (или однозначность) – каждому кодону соответствует только одна аминокислота. Вырожденность – одной аминокислоте может соответствовать несколько кодонов. Универсальность – биологический код одинаков для всех видов организмов на Земле (однако в митохондриях млекопитающих есть исключения). Колинеарность – последовательность кодонов соответствует последовательности аминокислот в кодируемом белке. Неперекрываемость – триплеты не накладываются друг на друга, располагаясь рядом. Отсутствие знаков препинания – между триплетами нет дополнительных нуклеотидов или каких-либо иных сигналов. Однонаправленность – при синтезе белка считывание кодонов идет последовательно, без пропусков или возвратов назад. Однако ясно, что биологический код не может проявить себя без дополнительных молекул, которые выполняют переходную функцию или функцию адаптора. А 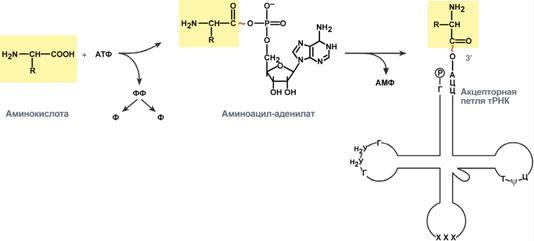 дапторная роль транспортных РНК. Транспортные РНК являются единственным посредником между 4-х буквенной последовательностью нуклеиновых кислот и 20-ти буквенной последовательностью белков. Каждая транспортная РНК имеет определенную триплетную последовательность в антикодоновой петле (антикодон) и может присоединить только такую аминокислоту, которая соответствует этому антикодону. Именно от наличия того или иного антикодона в тРНК зависит, какая аминокислота включится в белковую молекулу, т.к. ни рибосома, ни мРНК не узнают аминокислоту. Таким образом, адапторная роль тРНК заключается: дапторная роль транспортных РНК. Транспортные РНК являются единственным посредником между 4-х буквенной последовательностью нуклеиновых кислот и 20-ти буквенной последовательностью белков. Каждая транспортная РНК имеет определенную триплетную последовательность в антикодоновой петле (антикодон) и может присоединить только такую аминокислоту, которая соответствует этому антикодону. Именно от наличия того или иного антикодона в тРНК зависит, какая аминокислота включится в белковую молекулу, т.к. ни рибосома, ни мРНК не узнают аминокислоту. Таким образом, адапторная роль тРНК заключается:1.в специфичном связывании с аминокислотами, 2.в специфичном, согласно кодон-антикодоновому взаимодействию, связывании с мРНК, 3.и, как результат, во включении аминокислот в белковую цепь в соответствии с информацией мРНК. Присоединение аминокислоты к тРНК осуществляется ферментом аминоацил-тРНК-синтетазой, имеющей специфичность одновременно к двум соединениям: какой-либо аминокислоте и соответствующей ей тРНК. Для реакции требуется две макроэргические связи АТФ. Аминокислота присоединяется к 3'-концу акцепторной петли тРНК через свою α-карбоксильную группу, и связь между аминокислотой и тРНК становится макроэргической. α-Аминогруппа остается свободной. Так как существует около 60 различных тРНК, то некоторым аминокислотам соответствует по две или более тРНК. Различные тРНК, присоединяющие одну аминокислоту, называют изоакцепторными. Т 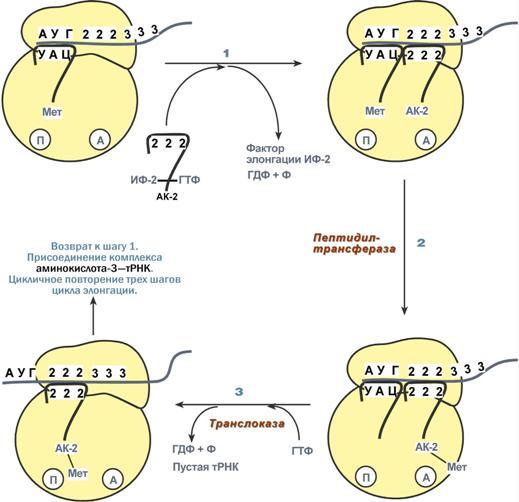 рансляция – это биосинтез белка на матрице мРНК. рансляция – это биосинтез белка на матрице мРНК.После переноса информации с ДНК на матричную РНК начинается синтез белков. Каждая зрелая мРНК несет информацию только об одной полипептидной цепи. Если клетке необходимы другие белки, то необходимо транскрибировать мРНК с иных участков ДНК. Биосинтез белков или трансляция происходит на рибосомах, внутриклеточных белоксинтезирующих органеллах, и включает 5 ключевых элементов: 1)матрица – матричная РНК, 2)растущая цепь – полипептид, 3)субстрат для синтеза – 20 протеиногенных аминокислот, 4)источник энергии – ГТФ, 5)рибосомальные белки, рРНК и белковые факторы. Выделяют три основных стадии трансляции: инициация, элонгация, терминация. И 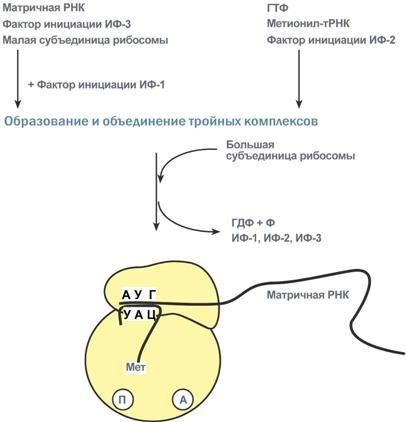 нициация. Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых фактора инициации (ИФ-1, ИФ-2, ИФ-3), метионин и тРНК для метионина. нициация. Для инициации необходимы мРНК, ГТФ, малая и большая субъединицы рибосомы, три белковых фактора инициации (ИФ-1, ИФ-2, ИФ-3), метионин и тРНК для метионина.В начале этой стадии формируются два тройных комплекса: первый комплекс – мРНК + малая субъединица + ИФ-3, второй комплекс – метионил-тРНК + ИФ-2 + ГТФ. После формирования тройные комплексы объединяются с большой субъединицей рибосомы. В этом процессе активно участвуют белковые факторы инициации, источником энергии служит ГТФ. После сборки комплекса инициирующая метионил-тРНК связывается с первым кодоном АУГ матричной РНК и располагается в П-центре (пептидильный центр) большой субъединицы. А-центр (аминоацильный центр) остается свободным, он будет задействован на стадии элонгации для связывания аминоацил-тРНК. После присоединения большой субъединицы начинается стадия элонгации. Элонгация. Для этой стадии необходимы все 20 аминокислот, тРНК для всех аминокислот, белковые факторы элонгации, ГТФ. Удлинение цепи происходит со скоростью примерно 20 аминокислот в секунду. Элонгация представляет собой циклический процесс. Первый цикл (и следующие циклы) элонгации включает три шага: 1.Присоединение аминоацил-тРНК (еще второй) к кодону мРНК (еще второму), аминокислота при этом встраивается в А-центр рибосомы. Источником энергии служит ГТФ. 2.Фермент пептидилтрансфераза осуществляет перенос метионина с метионил-тРНК (в П-центре) на вторую аминоацил-тРНК (в А-центре) с образованием пептидной связи между метионином и второй аминокислотой. При этом уже активированная СООН-группа метионина связывается со свободной NH2-группой второй аминокислоты. Здесь источником энергии служит макроэргическая связь между аминокислотой и тРНК. 3.Фермент транслоказа перемещает мРНК относительно рибосомы таким образом, что первый кодон АУГ оказывается вне рибосомы, второй кодон (на рисунке ) становится напротив П-центра, напротив А-центра оказывается третий кодон (на рисунке ). Для этих процессов необходима затрата энергии ГТФ. Так как вместе с мРНК перемещаются закрепленные на ней тРНК, то инициирующая первая тРНК выходит из рибосомы, вторая тРНК с дипептидом помещается в П-центр. Второе повторение цикла – начинается с присоединения третьей аминоацил-тРНК к третьему кодону мРНК, аминокислота-3 становится в А-центр. Далее трансферазная реакции повторяется и образуется трипептид, занимающий А?центр, после чего он смещается в П-центр в транслоказной реакции. В пустой А-центр входит четвертая аминоацил-тРНК и начинается третий цикл элонгации: Ц 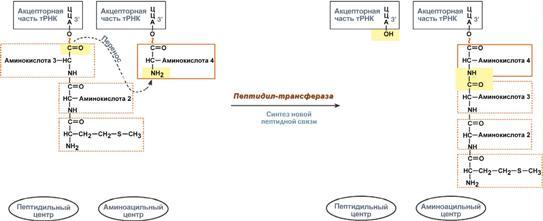 икл элонгации (реакции 1,2,3) повторяется столько раз, сколько аминокислот необходимо включить в полипептидную цепь. икл элонгации (реакции 1,2,3) повторяется столько раз, сколько аминокислот необходимо включить в полипептидную цепь.Т 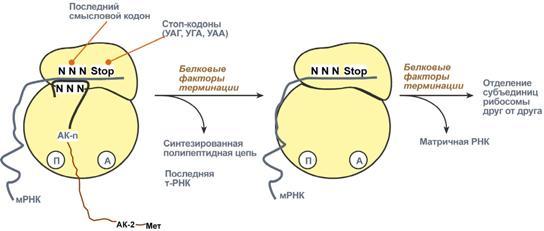 ерминация. Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК особых терминирующих кодонов – стоп-кодонов УАА, УАГ, УГА. Данные триплеты не кодируют ни одной из аминокислот, их также называют нонсенс-кодоны. При вхождении этих кодонов внутрь рибосомы происходит активация белковых факторов терминации, которые последовательно катализируют: ерминация. Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК особых терминирующих кодонов – стоп-кодонов УАА, УАГ, УГА. Данные триплеты не кодируют ни одной из аминокислот, их также называют нонсенс-кодоны. При вхождении этих кодонов внутрь рибосомы происходит активация белковых факторов терминации, которые последовательно катализируют:1)Гидролитическое отщепление полипептида от конечной тРНК. 2)Отделение от П-центра последней, уже пустой, тРНК. 3)Диссоциацию рибосомы. Источником энергии для завершения трансляции является ГТФ. По причине того, что продолжительность жизни матричной РНК невелика, перед клеткой стоит задача использовать ее максимально эффективно, т.е. получить максимальное количество "белковых копий". Для достижения этой цели на каждой мРНК может располагаться не одна, а несколько рибосом, встающих последовательно друг за другом и синтезирующих пептидные цепи. Такие образования называются полирибосомы. После того как пептидная цепь отходит от рибосомы она должна принять свою биологически активную форму, т.е. свернуться определенным образом, связать какие-либо группы и т.п. Реакции превращения полипептида в активный белок называются процессинг или посттрансляционная модификация белков. Посттрансляционная модификация белков. К основным реакциям процессинга относятся: 1. Удаление с N-конца метионина или даже нескольких аминокислот специфичными аминопептидазами. 2. Образование дисульфидных мостиков между остатками цистеина. 3. Частичный протеолиз – удаление части пептидной цепи, как в случае с инсулином или протеолитическими ферментами ЖКТ. 4. Присоединение химической группы к аминокислотным остаткам белковой цепи: фосфорной кислоты – например, фосфорилирование по аминокислотам Серину, Треонину, Тирозину используется при регуляции активности ферментов или для связывания ионов кальция, карбоксильной группы – например, при участии витамина К происходит γ-карбоксилирование глутамата в составе протромбина, проконвертина, фактора Стюарта, Кристмаса, что позволяет связывать ионы кальция при инициации свертывания крови, метильной группы – например, метилирование аргинина и лизина в составе гистонов используется для регуляции активности генома, гидроксильной группы – например, образование гидроксипролина и гидроксилизина необходимо для созревания молекул коллагена при участии витамина С, йода – например, в тиреоглобулине присоединение йода необходимо для образования предшественников тиреоидных гормонов йодтиронинов, 5. Включение простетической группы: углеводных остатков – например, гликирование требуется при синтезе гликопротеинов. гема – например, при синтезе гемоглобина, миоглобина, цитохромов, каталазы, витаминных коферментов – биотина, ФАД, пиридоксальфосфата и т.п. 6. Объединение протомеров в единый олигомерный белок, например, гемоглобин, коллаген, лактатдегидрогеназа, креатинкиназа. Фолдинг белков – это процесс укладки вытянутой полипептидной цепи в правильную трехмерную пространственную структуру. Для обеспечения фолдинга используется группа вспомогательных белков под названием шапероны (chaperon, франц. – спутник, нянька). Они предотвращают взаимодействие новосинтезированных белков друг с другом, изолируют гидрофобные участки белков от цитоплазмы и "убирают" их внутрь молекулы, правильно располагают белковые домены. В целом шапероны способствуют переходу структуры белков от первичного уровня до третичного и четвертичного. При нарушении функции шаперонов и отсутствии фолдинга в клетке формируются белковые отложения – развивается амилоидоз. Насчитывают около 15 вариантов амилоидоза. 41. Регуляция биосинтеза белка. Индукция и репрессия синтеза белка на примере функционирования лактозного оперона кишечной палочки. Ингибиторы матричных биосинтезов: лекарственные препараты, вирусные и бактериальные токсины. Так как транскрипция связывает ядро – "мозг" клетки, ее "банк знаний" и белки, "рабочих лошадок" клетки, то от качества и активности транскрипции зависит объем синтеза тех или иных белков, жизнедеятельность клетки, ее способность адаптироваться к окружающей обстановке.У прокариот и эукариот регуляция транскрипции происходит, естественно, по-разному, хотя некоторые моменты похожи. Регуляция биосинтеза белка у прокариот осуществляется на уровне изменения скорости синтеза мРНК. В настоящее время принята теория оперона, сформулированная Франсуа Жакобом и Жаком Моно. В основе теории лежат следующие понятия: 1 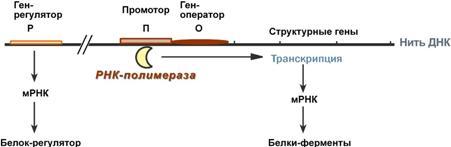 .оперон – группа тесно связанных между собой генов, которые программируют образование структурных белков и ферментов в клетке, .оперон – группа тесно связанных между собой генов, которые программируют образование структурных белков и ферментов в клетке,2.конституитивные ферменты – те, которые присутствуют в клетках всегда, независимо от ее активности и условий, 3.индуцибельные ферменты – те, которые программируются опероном и синтезируются при необходимости, 4.ген-регулятор – ген, регулирующий работу оперона, но не входящий в его состав. Он синтезирует белок-регулятор (чаще называемый белок-репрессор), который может быть в активной или неактивной форме, 5.ген-операто –участок ДНК, способный св-ся с белком-регулятором, и "решающий" нужно работать РНК-полимеразе или нет. П 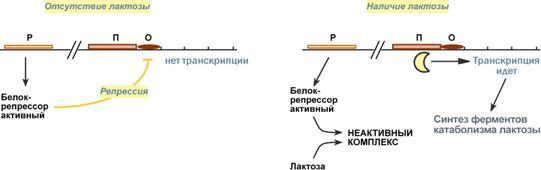 редложены две схемы регуляции скорости транскрипции: по механизму индукции (лактозный оперон) и по механизму репресии (триптофановый оперон). Лактозный оперон в целом отвечает за катаболизм лактозы. При изучении E.coli было замечено, что в клетке может быть две взаимоисключающие ситуации: редложены две схемы регуляции скорости транскрипции: по механизму индукции (лактозный оперон) и по механизму репресии (триптофановый оперон). Лактозный оперон в целом отвечает за катаболизм лактозы. При изучении E.coli было замечено, что в клетке может быть две взаимоисключающие ситуации:-активность одного из ферментов катаболизма лактозы низка, если в среде имеется глюкоза. -активность этого фермента резко повышается в обратной ситуации, т.е. при отсутствии глюкозы и при наличии лактозы.На основании наблюдений была предложена схема регуляции оперона по механизму индукции: 1. В отсутствие лактозы активный белок-репрессор связывается с оператором и блокирует синтез мРНК, кодирующей ферменты катаболизма лактозы. В результате эти ферменты не образуются. 2. Если глюкозы нет, а лактоза есть, то последняя связывается с белком-репрессором и ингибирует его, не давая связаться с геном-оператором. Это позволяет РНК-полимеразе считывать информацию, отвечающую за синтез ферментов катаболизма лактозы, и синтезировать мРНК. Т.о., лактоза является индуктором транскрипции. Триптофановый оперон в целом отвечает за синтез триптофана.Функционирование триптофанового оперона в некотором смысле противоположно лактозному. Регуляция осуществляется по механизму репрессии. 1. В отличие от лактозного оперона, белок-репрессор синтезируется в неактивном состоянии и не может заблокировать транскрипцию генов, кодирующих ферменты синтеза триптофана. Синтез этой аминокислоты будет в клетке продолжаться до тех пор, пока в питательной среде не появится триптофан. 2. Триптофан соединяется с белком-репрессором и активирует его. Далее такой активный комплекс присоединяется к гену-оператору и блокирует транскрипцию. Таким образом, при наличии триптофана в среде прекращается его внутриклеточный синтез, экономятся ресурсы и энергия бактериальной клетки. В этом случае триптофан является репрессором транскрипции. |