Ответы на билеты биохимия 2012. Белок это последовательность ак, связанных друг с другом пептидными связями
 Скачать 5.15 Mb. Скачать 5.15 Mb.
|

|
Креатин образуется в последовательных реакциях в почках и печени и далее доставляется в скелетные мышцы, миокард и нервную ткань. Здесь он фосфорилируется и выполняет роль резервного макроэрга. В норме в моче нет. Клиникодиагностическое значение. В первые годы жизни ребенка возможна физиологическая креатинурия, что объясняется его усиленным синтезом, опережающим рост мускулатуры. Креатинурия возможна и в пожилом возрасте как следствие атрофии мышц и снижения использования образующегося в печени креатина. У взрослых увеличение содержания креатина в крови свыше 0,12 ммоль/л сопровождается появлением его в моче. Выделение креатина возрастает при беременности и в раннем послеродовом периоде. Накопление в моче отмечается при поражениях мышечной системы (миопатии, мышечная дистрофия), при сахарном диабете, эндокринных расстройствах (гипертиреоз, аддисонова болезнь, акромегалия), инфекционных заболеваниях, системной красной волчанке, переломах костей, ожогах, белковом голодании, недостатке витамина Е. Креатинфосфокиназа (Креатинкиназа), магнийзависимый фермент, содержится исключительно в цитоплазме и митохондриях миокарда, скелетной мускулатуры и ткани мозга, где катализирует реакцию: Креатин + АТФ АДФ + Креатинфосфат Равновесие реакции при щелочных значениях pH сдвинуто в сторону образования АТФ. Высокая активность фермента обнаружена в мышечной и нервной ткани, в которых значительная часть энергии переносится между клеточными компартментами в виде креатинфосфата. Среди различных типов мышечной ткани активность КФК распределяется следующим образом: поперечно полосатые мышцы > сердечные мышцы > мышцы беременной матки > мышцы матки > гладкие мышцы. Фермент является гетерогенным белком, состоящим из 2 х типов субъединиц — В и М. В связи с этим выделяют три изофермента: ММ (содержится в скелетной мускулатуре и миокарде), ВВ (преимущественно в мозге и гладких мышцах) и МВ (в сердечной мышце). Изоферменты различаются по физико химическим и иммунологическим свойствам. Наиболее широкое применение исследование активности КФК нашло при диагностике инфаркта миокарда: активность фермента возрастает уже через 2 3 часа после поражения миокарда и через 14 30 часов достигает максимума, нормализация показателей наступает обычно на вторые-третьи сутки после приступа, длительное удерживание гиперэнзимемии наблюдается у больных с выраженным нарушением коронарного кровообращения. Кроме инфаркта, увеличение активности фермента в сыворотке стимулируют миокардиты, сердечная недостаточнсть, сердечные аритмии, при этом отмечается 20 30 кратное превышение нормальных показателей. Увеличение идет в основном за счет MB фракции КФК. Значительно более высокая активность КФК обнаруживается при заболеваниях мышечной системы, прежде всего миогенного происхождения (полиомиелит, мышечная дистрофия, дерматомиозит). При прогрессирующей мышечной дистрофии (миопатии) увеличение активности КФК отмечается уже в первые стадии болезни. В конечной стадии, вследствие замещения мышечной ткани на соединительную и жировую, активность фермента приходит в норму. Подъем активности может наблюдаться при травматическом повреждении ткани мозга, инфаркте мозга, менингите, гипотиреозе, отравлениях снотворными, кровоизлияниях в мозг. Снижение активности не имеет практического значение, отражая либо малую мышечную массу, либо сидячий образ жизни. 35. Нуклеозиды, нуклеотиды и нуклеиновые кислоты, строение, значение. Отличия ДНК и РНК. Нуклеопротеиды. Переваривание нуклеопротеидов. Нуклеиновые кислоты — высокомолекулярные соединения со строго определенной линейной последовательностью мономеров (мононуклеотидов), носители генетической информации обо всех белках, работающих в организме. В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). В состав нуклеиновых кислот входят азотистые основания двух типов: пуриновые - аденин (А), гуанин (G) и пиримидиновые - цитозин (С), тимин (Т) и урацил (U). Пентозы в нуклеотидах представлены либо рибозой (в составе РНК), либо дезоксирибозой (в составе ДНК). Пентозу соединяет с основанием N-гликозидная связь, образованная С1-атомом пентозы (рибозы или дезоксирибозы) и N1 -атомом пиримидина или N9-aтомом пурина. Нуклеозиды — это гликозиламины, содержащие азотистое основание, связанное с сахаром (рибозой или дезоксирибозой). Нуклеоти́ды — фосфорные эфиры нуклеозидов, нуклеозидфосфаты. Мономерные единицы из которых состоит ДНК и РНК. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов. Нуклеиновые кислоты по своему строению относят к классу линейных полимеров. Остов нуклеиновой кислоты имеет одинаковое строение по всей длине молекулы и состоит из чередующихся групп - пентоза-фосфат-пентоза- . Вариабельными группами в полинуклеотидных цепях служат азотистые основания - пурины и пиримидины. В РНК входят (А), (U), (G) и (С), в ДНК - (А), (Т), (G) и (С). Первичная структура ДНК - порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинукпеотидной цепи. Каждая фосфатная группа в полинукпеотидной цепи, за исключением фосфорного остатка на 5'-конце молекулы, участвует в образовании двух эфирных связей с участием 3'- и 5'-углеродных атомов двух соседних дезоксирибоз, поэтому связь между мономерами обозначают 3', 5'-фосфодиэфирной. Концевые нуклеотиды ДНК различают по структуре: на 5'-конце находится фосфатная группа, а на 3'-конце цепи - свободная ОН-группа. Эти концы называют 5'- и 3'-концами. Азотистые основания почти нерастворимы в воде, но некоторые атомы пуринового и пиримидинового циклов способны образовывать водородные связи. Вторичная структура - Двойная спираль правозакрученная, полинуклеотидньхе цепи в ней антипараллельны. Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов - снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счёт водородных связей между комплементарными пуриновыми и пиримидиновыми азотистыми основаниями А и Т (две связи) и между G и С (три связи). Правило Чаргаффа: « число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С)». Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль. В образованной структуре различают две бороздки – большую и малую. Азотистые основания в их области взаимодействуют со специфическими белками, участвующими в организации структуры хроматина. Нуклеопротеиды — комплексы нуклеиновых кислот с белками., а хроматин относятся как раз к этим комплексам (Кроме этого разнообразные вирусные нуклеопротеиды). Каждая молекула ДНК упакована в отдельную хромосому. Компактизация и суперспирализация ДНК осуществляются с помощью гистоновых белков. Гистоны - белки содержащие много остатков аргинина и лизина. Благодаря положительному заряду гистоны образуют ионные связи с отрицательно заряженными фосфатными группами, расположенными на внешней стороне двойной спирали ДНК. Молекула ДНК "накручивается" на поверхность гистонового октамера. акой комплекс гистоновых белков с ДНК служит основной структурной единицей хроматина, её называют "нуклеосома". Первичная структура РНК - порядок чередования рибонуклеозидмонофосфатов (НМФ) в полинуклеотидной цепи. В РНК, как и в ДНК, нук-леотиды связаны между собой 3',5'-фосфодиэфирными связями. Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная ОН-группа 5'-углеродного атома, на другом конце - ОН-группа 3'-углеродного атома рибозы, поэтому концы называют 5'- и 3'-концами цепи РНК. Гидроксильная группа у 2'-углеродного атома рибозы делает молекулу РНК нестабильной. Так, в слабощелочной среде молекулы РНК гидролизуются даже при нормальной температуре, тогда как структура цепи ДНК не изменяется. Вторичная структура РНК - Молекула рибонуклеиновой кислоты построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли - "шпильки", за счёт водородных связей между комплементарными азотистыми основаниями A-U и G-C. Участки цепи РНК в таких спиральных структурах антипараллельны, но не всегда полностью комплементарны, в них встречаются неспаренные нуклеотидные остатки или даже одноцепочечные петли, не вписывающиеся в двойную спираль. Наличие спирализованных участков характерно для всех типов РНК. Одноцепочечные РНК характеризуются компактной и упорядоченной третичной структурой, возникающей путём взаимодействия спирализованных элементов вторичной структуры. Так, возможно образование дополнительных водородных связей между нуклеотидными остатками, достаточно удалёнными друг от друга, или связей между ОН-группами остатков рибо-зы и основаниями. Третичная структура РНК стабилизирована ионами двухвалентных металлов, например ионами Mg2+, связывающимися не только с фосфатными группами, но и с основаниями. В цитоплазме клеток присутствуют 3 типа рибонуклеиновых кислот - транспортные РНК (тРНК), матричные РНК (мРНК) и рибосомальные РНК (рРНК). Они различаются по первичной структуре, молекулярной массе, конфор-мации, продолжительности жизни и, самое главное, по функциональной активности. Отличия РНК и ДНК.1)Молекулярная масса РНК 25кД, ДНК от 1000 до 1000000 кД. 2)Моносахарид (пентоза) в РНК представлен рибозой, в ДНК дезоксирибозой. 3)Азотистые основания в РНК- аденин, урацил, гуанин, цитозин; в ДНК- аденин, Тимин, гуанин, цитозин. 4)Первичная структура РНК нестабильна, в отличии от ДНК, т.к. имеет гидроксильную группу у 2`- углеводного атома рибозы. Переваривание нуклеопротеидов. Нуклеиновый компонент отделяется от белка в кислой среде желудка. Катаболизм нуклеиновых кислот начинается с гидролиза 3',5'-фосфодиэфирной связи под действием ферментов нуклеаз. 1. ДНКазы - расщепляют ДНК. 2. РНКазы - расщепляют РНК. Среди ДНКаз и РНКаз различают: 1) экзонуклеазы (5' и 3'); 2) эндонуклеазы - специфичны к мононуклеотидной последовательности, есть высокоспецифичные: рестриктазы - используются в генной инженерии. Далее происходит отщепление фосфата от мононуклеотида с участием ферментов нуклеотидаз с образованием нуклеозидов. Нуклеозид может расщепляться путем гидролиза под действием фермента нуклеозидазы на азотистое основание и пентозу, но чаще происходит фосфоролиз - при этом нуклеозид расщепляется на азотистое основание и фосфорибозу. Продукты гидролиза полинуклеотидов — мононуклеотиды под действием нуклеотидаз и нуклеозидаз расщепляются на азотистые основания, пентозы (рибоза или дезоксирибоза) и фосфорную кислоту, которые всасываются. Нуклеазы очень многообразны. Различают ДНКазы и РНКазы. Отличаются друг от друга по месту действия. Бывают эндонуклеазы (расщепляют внутренние 3'5'-фосфодиэфирные связи) и экзонуклеазы (отщепляют концевые мононуклеотиды). Встречается 2 типа экзонуклеаз: 3'-экзонуклеазы - отщепляют мононуклеотид с 3'-конца молекулы, и 5'-экзонуклеазы - отщепляют 5'-концевой мононуклеотид. Нуклеазы могут отличаться друг от друга и по специфичности. Пищеварительные нуклеазы и нуклеазы лизосомальные отличаются низкой специфичностью, у них упрощенное строение активного центра. У высокоспецифичных нуклеаз очень сложное строение активного центра. Они способны "узнавать" целую последовательность нуклеотидов из 4-10 пар и расщеплять одну-единственную связь в строго определенном месте. Такие высокоспецифичные ДНКазы называются рестриктазами. Участки ДНК, "узнаваемые" рестриктазами, называются полиндромными последовательностями. Пентозы, образующиеся в ходе катаболизма нуклеиновых кислот, могут быть утилизированы во II-м этапе ГМФ-пути. Азотистые основания также подвергаются дальнейшему катаболизму, но по-разному, в зависимости от их типа - пуриновых (аденина, гуанина) или пиримидиновых (тимина, цитозина и урацила). 36. Катаболизм пуриновых и пиримидиновых оснований. Гиперурикемия. Подагра. Катаболизм пуриновых оснований. Распад начинается с отщепления. От АМФ и аденозина аминогруппа удаляется гидролитически аденозиндезаминазой с образованием ИМФ или инозина. ИМФ и ГМФ превращаются в соответствующие нуклеозиды: инозин и гуанозин под действием 5´-нуклеотидазы. Пуриннуклеозидфосфорилаза катализирует расщепление N-гликозидной связи в инозине и гуанозине с образованием рибозо-1-фосфата и азотистых оснований: гуанина и гипоксантина. Гуанин дезаминируется и превращается в ксантин, а гипоксантин окисляется в ксантин с помощью ксантиноксидазы, которая катализирует и дальнейшее окисление ксантина в мочевую кислоту. Образовавшийся инозин подвергается фосфоролизу, и далее гипоксантин подвергается двукратному окислению путем отнятия водорода с одновременным присоединением воды. Эти две одинаковые реакции катализирует один и тот же фермент - ксантиноксидаза. Ксантиноксидаза - аэробная оксидоредуктаза.Она может существовать в двух формах:1) D-форма - дегидрогеназная; 2) O-форма - оксидазная. Формы отличаются друг от друга по способности передавать 2 атома водорода. D-форма передает водород на главную дыхательную цепь митохондриального окисления, а O-форма - сразу на кислород с образованием H2O2. D-форма может переходить в О-форму путем ограниченного протеолиза при отщеплении небольшого участка молекулы. Образование мочевой кислоты происходит главным образом в печени. Мочевая кислота — основной продукт катаболизма пуриновых нуклеотидов у человека. В организме ежесуточно обра¬зуется 0,5 — 1 г мочевой кислоты, которая выводится через почки. Мочевая кислота плохо растворима в воде и жидких средах организма. В норме ее концентрация на пределе растворимости и составляет 0,1 — 0,4 ммоль/л. Если концентрация мочевой кислоты становится выше нормальной, происходит отложение ее солей (уратов) в суставах и почках. Повышение концентрации мочевой кислоты в крови называется гиперурикемией. Однако повышенное содержание мочевой кислоты в крови (гиперурикемия) небезопасно. Сама мочевая кислота и, особенно, ее соли ураты (натриевые соли мочевой кислоты) плохо растворимы в воде. Даже при незначительном повышении концентрации они начинают начинают выпадать в осадок и кристаллизоваться, образуя камни. Кристаллы воспринимаются организмом как чужеродный объект. В суставах они фагоцитируются макрофагами, сами клетки при этом разрушаются, из них освобождаются гидролитические ферменты. Это приводит к воспалительной реакции, сопровождающейся сильнейшими болями в суставах. Такое заболевание называется подагра. Другое заболевание, при котором кристаллы уратов откладываются в почечной лоханке или в мочевом пузыре, известно как мочекаменная болезнь. Классическая подагра обусловлена тремя факторами — увеличенным синтезом мочевой кислоты, снижением содер¬жания в плазме уратсвязывающего белка и замедленным выведением с мочой. Клинические проявления подагры: 1 Повторяющиеся приступы острого воспаления суставов, чаще всего мелких (подагрический криз); обычно болезнь начинается с воспаления первого плюснефалангового сустава большого пальца ноги. Подагрический криз связан с отложением кристаллов мононатриевой соли мочевой кислоты в суставе. 2. Подагрические узлы (тофусы), возникают в результате местного отложения и накопления уратов; локализация — мелкие суставы, сухожилия, хрящи, кожа. Для лечения подагры применяются: 1) ингибиторы фермента ксантиноксидазы. Например, аллопуринол - вещество пуриновой природы, является конкурентным ингибитором фермента. Действие этого препарата приводит к повышению концентрации гипоксантина. Гипоксантин и его соли лучше растворимы в воде, и легче выводятся из организма.2) диетическое питание, исключающее продукты, богатые нуклеиновыми кислотами, пуринами и их аналогами: икра рыб, печень, мясо, кофе и чай. 3) соли лития, поскольку они лучше растворимы в воде, чем ураты натрия. Катаболизм пиримидиновых нуклеотидов. Превращения пиримидиновых оснований в результате деза-минирования и декарбоксилирования идет до (3-аланина, угле¬кислого газа, аммиака и (3-аминоизомасляной кислоты. Пиримидиновые основания при участии дигидропиримидиндегидрогеназы присоединяют 2 атома водорода по двойной связи кольца с образованием дигидроурацила или дигидротимина. Оба гетероцикла могут взаимодействовать с водой в реакции, катализируемой дигидропиримидинциклогидролазой, и дигидроурацил превращается в β-уреидопропионовую кислоту, а дигидротимин - в β-уреидоизомасляную кислоту. Оба β-уреидопроизводных под действием общего для них фермента уреидопропионазы расщепляются с образованием СО2, NH4+ и β-аланина или β-аминоизомасляной кислоты соответственно. β-Аланин обнаруживают в плазме крови и многих тканях. Он используется в мышцах на образование дипептидов: карнозина и анзерина. Под действием бактериальной микрофлоры кишечника β-аланин включается в пантотеновую кислоту, которая всасывается и используется на образование КоА. Часть β-аланина и β-аминбутирата трансами-нируется с α-кетоглутаратом и даёт малонил полуальдегид или метилмалонил полуальдегид соответственно, которые превращаются в малонил-КоА и сукцинил-КоА и используются в соответствующих метаболических путях, либо окисляются до СО2 и Н2О. Частично β-аминобутират экскретируется с мочой. 37. Биосинтез пуриновых и пиримидиновых нуклеотидов. Биосинтез дезоксирибонуклеотидов. Регуляция этих процессов. Синтез пуриновых оснований происходит во всех клетках организма, главным образом в печени. Исключение составляют эритроциты, полиморфноядерные лейкоциты, лимфоциты. Условно все реакции синтеза можно разделить на 4 этапа: 1 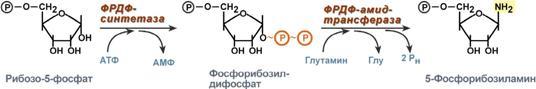 . Синтез 5'-фосфорибозиламина . Синтез 5'-фосфорибозиламинаПервая реакция синтеза пуринов заключается в активации углерода в положении С1 рибозо-5-фосфата, это достигается синтезом 5-фосфорибозил-1-дифосфата (ФРДФ). Рибозо-5-фосфат является тем якорем, на основе которого синтезируется сложный пуриновый цикл. Вторая реакция – это перенос NH2-группы глутамина на активированный атом С1 рибозо-5-фосфата с образованием 5'-фосфорибозиламина. Указанная NH2-группа фосфорибозиламина уже принадлежит будущему пуриновому кольцу и ее азот будет атомом номер 9. Параллельно фофорибозилдифосфат используется при синтезе пиримидиновых нуклеотидов. Он реагирует с оротовой кислотой и рибозо-5-фосфат связывается с ней, образуя оротидилмонофосфат (реакция). 2. Синтез инозинмонофосфата 5 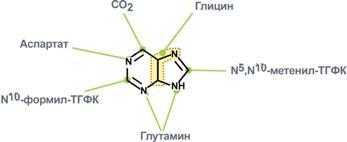 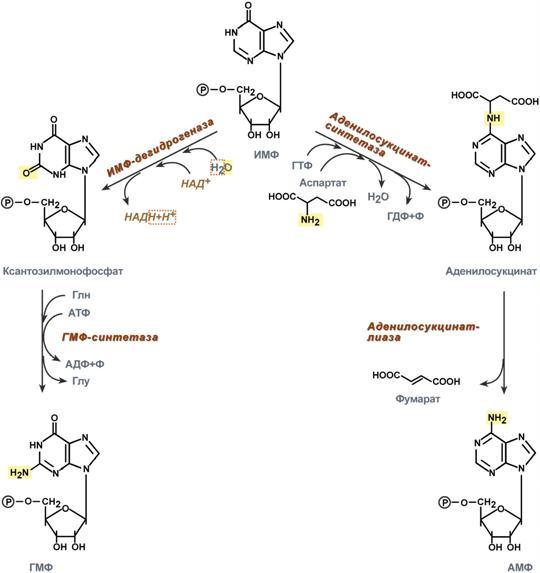 -фосфорибозиламин вовлекается в девять реакций, и в результате образуется первый пуриновый нуклеотид – инозинмонофосфорная кислота (ИМФ). В этих реакциях источниками атомов пуринового кольца являются глицин, аспартат, еще одна молекула глутамина, углекислый газ и производные тетрагидрофолиевой кислоты (ТГФК). В целом на синтез пуринового кольца затрачивается энергия 6 молекул АТФ. -фосфорибозиламин вовлекается в девять реакций, и в результате образуется первый пуриновый нуклеотид – инозинмонофосфорная кислота (ИМФ). В этих реакциях источниками атомов пуринового кольца являются глицин, аспартат, еще одна молекула глутамина, углекислый газ и производные тетрагидрофолиевой кислоты (ТГФК). В целом на синтез пуринового кольца затрачивается энергия 6 молекул АТФ.3. Синтез аденозинмонофосфата и гуанозинмонофосфата а) Гуанозинмонофосфат (ГМФ) образуется в двух реакциях – сначала он окисляется ИМФ-дегидрогеназой до ксантозилмонофосфата, источником кислорода является вода, акцептором водорода – НАД. После этого работает ГМФ-синтетаза, она использует универсальный клеточный донор NH2-групп – глутамин, источником энергии для реакции служит АТФ. б) Аденозинмонофосфат (АМФ) также образуется в двух реакциях, но в качестве донора NH2-группы выступает аспарагиновая кислота. В первой, аденилосукцинат-синтетазной, реакции на присоединение аспартата используется энергия распада ГТФ, во второй реакции аденилосукцинат-лиаза производит удаление части аспарагиновой кислоты в виде фумарата. 4. Образование нуклеозидтрифосфатов АТФ и ГТФ. С 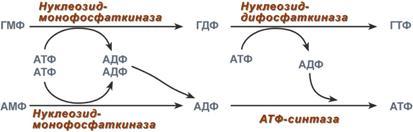 интез ГТФ осуществляется в 2 стадии посредством переноса макроэргических фосфатных групп от АТФ. Синтез АТФ происходит несколько иначе. АДФ из АМФ образуется также за счет макроэргических связей АТФ. Для синтеза же АТФ из АДФ в митохондриях есть фермент АТФ-синтаза, образующий АТФ в реакциях окислительного фосфорилирования. интез ГТФ осуществляется в 2 стадии посредством переноса макроэргических фосфатных групп от АТФ. Синтез АТФ происходит несколько иначе. АДФ из АМФ образуется также за счет макроэргических связей АТФ. Для синтеза же АТФ из АДФ в митохондриях есть фермент АТФ-синтаза, образующий АТФ в реакциях окислительного фосфорилирования. |