|
Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
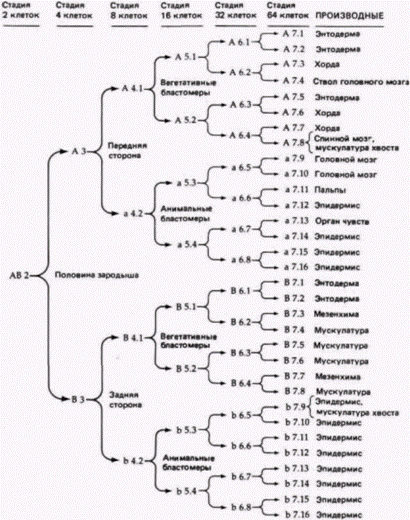
Рис. 7.3. Последовательность клеточных линии и их судьба в эмбриональном развитии оболочников. Поскольку правая и левая половины развиваются идентично, на рисунке представлена только одна половина. (По Whittaker. 1979; Meedel et al.. 1987; Nishida. 1987.)
|
|
|
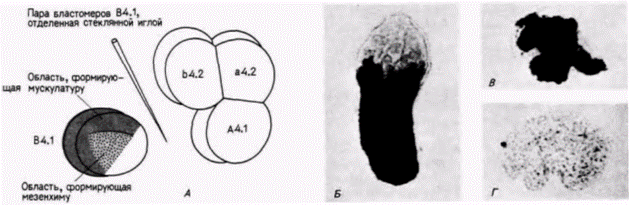
Рис. 7.4. Способность к синтезу ацетилхолинэстеразы в потомстве бластомеров мышечной линии (В4.1 ), изолированных на 8-клеточной стадии. А. Схема процедуры изоляции. Б. Локализация ацетилхолинэстеразы в хвостовой мускулатуре интактной личинки оболочника. Фермент обнаружен в потомстве пары бластомеров В4.1 (В), но не в потомстве остальных 6/8 зародыша (Г), инкубировавшихся в течение времени, необходимого для достижения стадии личинки. (Из Whittaker el al.. 1977; фотографии с любезного разрешения J. R. Whittaker.)
|
Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.
12________________ ГЛАВА 7______________________________________________________________________________

|
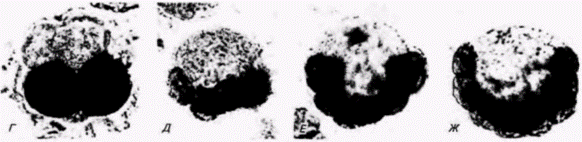
Рис. 7.5. Локализация ацетилхолинэстеразы в зародышах оболочников, развитие которых было остановлено на разных стадиях обработкой цитохалазином В. А. 1-клеточная стадия. Б. 2-клеточная стадия. В. 4-клеточная стадия. Г. 8-клеточная стадия. Д. 16-клеточная стадия. Е. 32-клеточная стадия. Ж. 64-клеточная стадия. (Из Whittakcr, 1973a; фотографии с любезного разрешения J. R. Wnittaker)
|
|
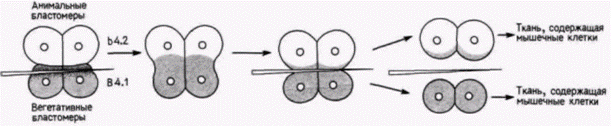
Рис. 7.6. Микрохирургическая процедура, позволяющая некоторому количеству цитоплазмы из области желтого серпа переместиться в анимальные клетки. В опыте использованы яйца оболочников. Надавливание на бластомеры B4.I стеклянной иглой вызывает регрессию борозды дробления. Борозда может образоваться вновь несколько ближе к вегетативному полюсу, там. где клетки были отрезаны иглой. Эта борозда отделяет пару бластомеров таким образом, что бластомеры анимального полюса (В4.2) получают цитоплазму желтого серпа. Цитоплазма вегетативного полюса на рисунке обозначена серым цветом. (Но Whittaker, 1982.)
|
В4.2. из него помимо нормальных эктодермальных клеток возникали еще и мышечные (рис. 7.6; Whittaker, 1982). Напротив. Тунг и др. (Tung et al., 1977) показали, что если ядра личинки трансплантировать в энуклеированные фрагменты яиц. то вновь образующиеся клетки имеют структуры, типичные для клеток - доноров цитоплазмы, а не для клеток, являющихся донорами ядер. Отсюда мы можем заключить, что определенные детерминанты, содержащиеся в цитоплазме, вызывают образование определенных тканей. Эти морфогенетические детерминанты (или морфогены), по-видимому, действуют посредством избирательной активации (или инактивации) специфических генов. Следовательно, детерминация бластомеров и активация определенных генов контролируются пространственной локализацией морфогенетических детерминантов в цитоплазме яйца.
Природа морфогенетических детерминантов у оболочников
Данные о природе цитоплазматических детерминантов противоречивы. Результаты исследований с применением ингибиторов транскрипции и транс-
|
|
|
 Скачать 19.05 Mb.
Скачать 19.05 Mb.