Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.__________________ ПРОГРЕССИВНАЯ ДЕТЕРМИНАЦИЯ_____________________________________________________ 41
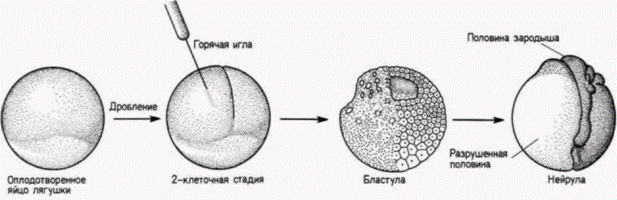
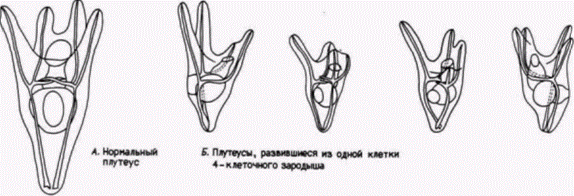
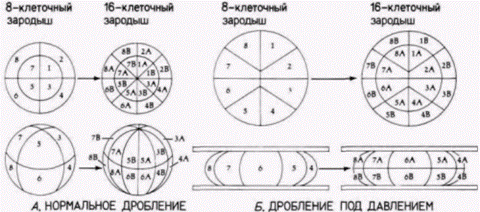
между двумя бластомерами. Это утверждение было проверено Вильгельмом Ру (W. Roux), молодым немецким эмбриологом, в прошлом учеником Эрнста Геккеля. Ру также был ревностным дарвинистом (в своей первой крупной работе по эмбриологии он высказал предположение о конкуренции между эмбриональными клетками), но, подобно Вейсману, чувствовал, что развитие должно изучаться с помощью аналитических методов. В 1888 г. Ру опубликовал результаты серии опытов, в которых он брал 2- и 4-клеточных зародышей лягушки и разрушал некоторые из клеток каждого зародыша горячей иглой. Из гипотезы Вейсмана следовало, что в этом случае образуется либо правая, либо левая половина зародыша. Ру и в самом деле получал половинные морулы, как то и предсказывал Вейсман (рис. 8.2). Развившиеся из них полунейрулы имели полный набор правой или левой стороны с одним нервным валиком, одной слуховой ямкой (плакодой) и т.д. На основе этих данных Ру пришел к выводу, что зародыш лягушки представляет собой мозаику самодифференцирующихся частей и очень вероятно, что каждая клетка получает свой набор детерминантов и соответственно этому дифференцируется. Этой серией опытов Ру начал свою программу по изучению механики развития (Entwicklungsmechanik), обосновывавшую физиологический подход к эмбриологии. Эмбриология. считал Ру, больше не должна быть просто служанкой эволюционных учений. Она должна выполнять свою роль независимой экспериментальной науки. Ганс Дриш: регуляционное развитиеНикто не приветствовал этот экспериментальный подход к эмбриологии больше, чем другой ученик Геккеля – Ганс Дриш (H. Driesch). Дриш ставил перед собой цель объяснить развитие с позиций законов физики и математики. Первые его исследования были сходны с исследованиями Ру. Методически опыты Ру были опытами с нанесением дефектов; они отвечали на вопрос, как будут развиваться оставшиеся бластомеры, когда часть их разрушена. Дриш (1892) задумал расширить эти исследования, проделав опыты с изоляцией бластомеров. Он отделял друг от друга бластомеры морского ежа энергичным встряхиванием (или позже помещением их в бескальциевую морскую воду). К удивлению Дриша, каждый из бластомеров двуклеточного зародыша развивался в полную личинку. Точно также, если Дриш разделял бластомеры 4и 8-клеточных зародышей, то некоторые из клеток образовывали целые личинки, называемые плутеусами (рис. 8.3). Этот результат поразительно отличался от того, что предсказывали Вейсман или Ру. Вместо самодифференцировки в будущую часть зародыша каждый бластомер мог регулировать свое развитие и давать начало целому организму. Это явление было названо регуляционным развитием. Регуляционное развитие было продемонстрировано Дришем и в других опытах. У морских ежей плоскости двух первых делений дробления являются меридиональными, тогда как третье деление проходит по экватору яйца, разделяя зародыш на четыре верхние и четыре нижние клетки (см. рис. 3.3). Дриш (1893) изменял направление третьего деления, слегка сжимая ранний зародыш между двумя стеклянными пластинками; в результате третье деление также проходило в меридиональной плоскости, как и первые два. Если затем давление ослабляли, то четвертое деление дробления было экваториальным. Эта процедура вызывала перегруппировку ядер, так что ядро, которое в норме должно было оказаться в области, предназначенной формировать энтодерму, теперь попадало в область презумптивной эктодермы. Ядра, которые должны были обра- Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.42________________ ГЛАВА 8_______________________________________________________________________________
зовывать дорсальные структуры, обнаруживались в вентральных клетках (рис. 8.4). Если бы в данном случае происходила сегрегация ядерных детерминантов, то у развившегося из этих клеток зародыша все структуры должны были располагаться в причудливом беспорядке. Однако Дриш получил из этих зародышей нормальных личинок. Результаты этих опытов оказались очень важными как для эмбриологии, так и лично для Дриша Во-первых. Дриш показал, что «проспективная потенция» изолированного бластомера (т.е. тот тип клеток, который мог из него произойти) шире, чем его «проспективная судьба» (т.е. те типы клеток, которые должны формироваться из этого бластомера при неизменном ходе его развития). Согласно же Вейсману и Ру. проспективная потенция и проспективная судьба бластомера должны быть идентичными. Во-вторых. Дриш пришел к выводу, что зародыш морского ежа представляет собой «гармоничную эквипотенциальную систему». Система эта гармонична потому, что все ее потенциально независимые части функционируют вместе, формируя единый организм. В-третьих, судьба ядра зависела исключительно от его положения в зародыше. Дриш (1894) выдвинул гипотезу о серии событий, продвигающих развитие вперед посредством взаимодействия ядра и цитоплазмы: «Поскольку каждая клетка содержит ядро, она на протяжении онтогенеза несет в себе всю сумму зачатков; поскольку она содержит специфическое
|