Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.226_____________ ГЛАВА 14___________________________________________________________________________
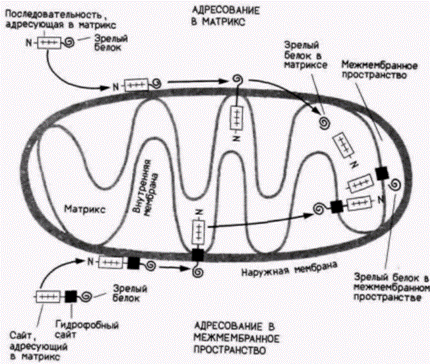
последовательность, состоящая из периодически расположенных основных аминокислот (конкретные длины и состав аминокислотных последовательностей у митохондриальных белков варьируют). Если эта последовательность находится в белке, то он не только направляется в митохондрии, но будет транслоцирован через обе мембраны в митохондриальный матрикс. Если между зрелым белком и адресующей в матрикс последовательностью лежит гидрофобный участок, то предполагается, что он блокирует перенос белка в матрикс. Это приведет к тому, что данный белок станет частью внутренней мембраны. Если зрелый белок легко отщепляется от этого гидрофобного участка, то он будет обнаруживаться в межмембранном пространстве (van Loon, Schatz, 1987). Надмолекулярная сборкаПоследняя форма посттрансляционной регуляции, которую мы рассмотрим, связана со сборкой новосинтезированных белков в функциональный комплекс. Мы уже рассмотрели ряд белков, которые могут спонтанно собираться в функциональные комплексы. Например, гемоглобин и лактатдегидрогеназа состоят из четырех полипептидных цепей, которые образуют с помощью нековалентных связей функциональный белок. Пример на несколько более высоком уровне – два сократительных белка, тубулин и актин, существуют в полимерных и неполимерных формах. Мы видели, что полимеризация актина из глобулярных мономеров в микрофила- Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.____________ ТРАНСЛЯЦИОННАЯ И ПОСТТРАНСЛЯЦИОННАЯ РЕГУЛЯЦИЯ РАЗВИТИЯ______________ 227
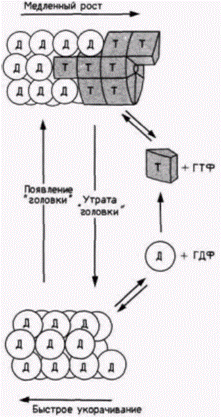
менты необходима для развития акросомного процесса при оплодотворении. Сходным образом тубулин собирается в микротрубочки митотического аппарата и распадается на мономеры при завершении митоза. Затем эти тубулиновые единицы могут быть реорганизованы в новые микротрубочки, которые важны для детерминации формы клеток. Рост микротрубочки регулируется гидролизом молекул ГТФ, которые ковалентно связаны с тубулином. Новополимеризованный тубулин присоединяется к ГТФ и образует стабильные комплексы с другими молекулами тубулина. При старении тубулинов (или при реорганизации клетки) ГТФ гидролизуется с образованием ГДФ. Тубулин, связанный с ГДФ, не образует комплекс с соседними молекулами тубулина. Полагают, что эта пониженная стабильность вызывает деполимеризацию микротрубочки (рис. 14.32). Как мы увидим в следующей главе, дифференциальный рост микротрубочек может оказывать определяющее влияние на клеточный и эмбриональный морфогенез. Способность формировать волокна свойственна |