Гилберт С. Биология развития. Т.2.doc ,БИР. Библиография Гилберт С. Биология развития в 3х т. Т. 2 Пер с англ. М. Мир, 1994. 235 с
 Скачать 19.05 Mb. Скачать 19.05 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.228_______________ ГЛАВА 14______________________________________________________________________________
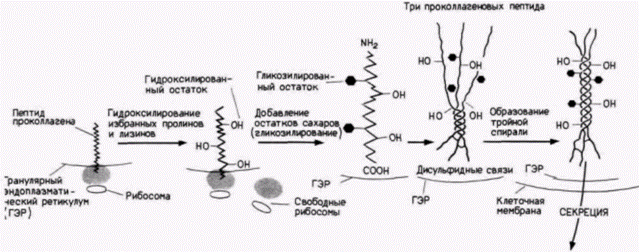
относительно небольшому набору клеточных белков. Приобретение этой способности другими белками благодаря мутации может приводить к гибельным последствиям. Такое явление наблюдается при серповидноклеточной анемии. Замена одной аминокислоты (глутамина на валин в шестом положении ß-цепи глобина) вызывает образование гидрофобного кармана, который позволяет взаимодействовать гемоглобиновым тетрамерам в восстановленной форме. Такие волокна растягивают клетку, придавая ей характерную форму серпа, и уменьшают ее гибкость (рис. 14.33). Некоторые белки образуют комплексы с нуклеиновыми кислотами. Например, гистоны нужны только для того, чтобы вместе с ДНК формировать нуклеосомные частицы. Эукариотическая 80S-рибосома состоит приблизительно из 70 белков и 4 различных рибосомных РНК. Эти белки и нуклеиновые кислоты для своего функционирования должны собираться вместе, и что примечательно, эта сборка происходит быстро и эффективно внутри всех клеток. После этого не должен вызывать удивление тот факт, что полноценные инфекционные вирусы могут быть получены при совместной инкубации структурных белков и нуклеиновых кислот. Белки, которые «маскируют» мРНК в ооцитах, также являются примерами белков, которые могут формировать специфические комплексы с нуклеиновыми кислотами. Белок TFIIIA, который комплексируется с 5S-pPHK в гене 5S-pPHK Xenopus, связывается также с 5S-pPHK в ооците и образует стабильную 7S-частицу (Pelham, Brown, 1980). Таким образом, спонтанная самосборка является еще одной важной формой посттрансляционного контроля. Коллаген: конспект посттрансляционной регуляцииПри рассмотрении синтеза коллагена, одного изглавнейших структурных белков тела, мы можем показать важность и разнообразие механизмов посттрансляционного контроля (см. обзоры Prockop et al., 1979; Davidson, Berg, 1981). Во-первых, последовательность мРНК коллагена транслируется в пептид проколлагена. Аминоконец проколлагена содержит сигнальную последовательность, которая позволяет ему войти в полость гранулярного эндоплазматического ретикулума. Этот сигнальный пептид отщепляется от остальной части молекулы коллагена. Внутри эндоплазмати- Гилберт С. Биология развития: В 3-х т. Т. 2: Пер. с англ. – М.: Мир, 1994. – 235 с.ТРАНСЛЯЦИОННАЯ И ПОСТТРАНСЛЯЦИОННАЯ РЕГУЛЯЦИЯ РАЗВИТИЯ___________ 229
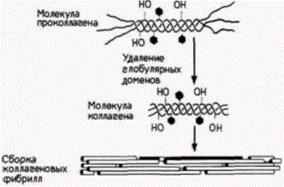
ческого ретикулума молекулы коллагена встречаются также с тремя ферментами, которые его гидроксилируют. Два из этих ферментов гидроксилируют остатки пролина в 3-гидроксипролин и 4-гидроксипролин; третий превращает лизины в оксилизиловые остатки. Эти ферменты узнают остатки пролина и лизина только в определенных положениях (рис. 14.34). Когда цепи коллагена становятся гидроксилированными, к оксилизинам могут быть добавлены остатки сахаров. Первый фермент – галактозил-трансфераза – добавляет к оксилизину галактозу; второй фермент – глюкозилтрансфераза – добавляет к оксилизилгалактозе глюкозу. Участок около карбоксильного конца модифицируется глюкозамином и маннозой. Следующий этап включает образование внутрицепочечных дисульфидных связей и организацию трех коллагеновых полипептидов в тройную спираль. Дисульфидные связи образуются на карбоксильных концах между прилежащими молекулами коллагена, создавая тем самым условия для формирования тройной спирали. Если пролины были гидроксилированы правильно, то три цепи скручиваются одна вокруг другой, образуя протяженный участок тройной спирали, ограниченный с обеих сторон глобулярными областями. В таком виде молекула коллагена выделяется во внеклеточное пространство. Процессинг продолжается даже там. Сначала протеолитические ферменты удаляют глобулярные домены на аминоконце и карбоксильном конце. В результате этого отрезания образуется фрагмент совершенной тройной спирали. На следующем этапе эти фрагменты самопроизвольно собираются в фибриллы. Однако эти фибриллы не обладают достаточной механической прочностью, чтобы функционировать в качестве структурных белков до тех пор, пока различные тройные спирали не будут ковалентно связаны друг с другом с помощью дополнительных поперечных сшивок. Это связывание осуществляется путем ферментативной модификации лизилов в оксилизиловых остатках и присоединения их друг к другу· Таким образом, существует значительное число необходимых этапов модификации белка после трансляции (как мы увидим в следующих двух главах, коллаген не только исключительно важный структурный белок, но, как полагают, он ответствен | ||||||