Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.________________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. МЕЗОДЕРМА И ЭНТОДЕРМА___________________________ 197
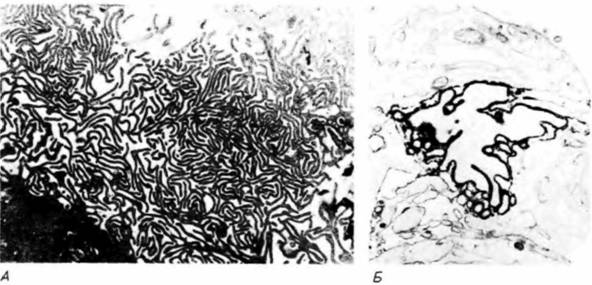
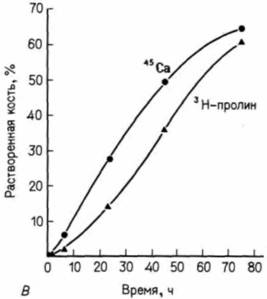
щается и митохондрии более не могут накапливать кальций (его накопление - активный энергоемкий процесс). В результат отмирающие хондроциты высвобождают кальций, и за его счет формируется провизорный обызвествленный (кальцифицированный) матрикс (рис. 6.12). Остеобласты выстраиваются в ряд вдоль обызвествленных полос матрикса и начинают откладывать на них кость. В конечном счете эти полосы исчезают и остается лишь кость, сформированная остеобластами. Таким образом, кость оказывается там, где ранее находился хрящ. Другая гипотеза подчеркивает различное происхождение начальных запасов фосфита кальция во внеклеточном матриксе. Согласно этой гипотезе, главными действующими агентами в накоплении кальция являются не митохондрии хондроцитов, а пузырьки матрикса. Считают, что эти пузырьки представляют собой специализированные участки плазматической мембраны хондроцита, которые отпочковываются от клетки и прилипают к внеклеточному матриксу. Попав на это место, они накапливают большие количества кальция и фосфита из окружающей лимфы (Dereszewski, Howell, 1978; Wuthier et al., 1985). Действительно, ли пузырьки связаны с самыми начальными отложениями кальция в хряще и кости. Недавно обнаружено (Wuthier et. al., 1985), что некоторые пузырьки содержат кальций из хондроцитов. Это позволяет предположить, что обе модели могут быть правильными и что накопление кальция может осуществляться с помощью не одного, а двух или более механизмов. Замещение хондроцитов остеобластами зависит, по-видимому, от минерализации внеклеточного матрикса. У куриных зародышей источником кальция Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.198 ГЛАВА 6 служит углекислый кальций скорлупы яйца. За время развития кровеносная система зародыша переносит около 120 мг кальция от скорлупы к скелету (Tuan, 1987). Если у развивающегося куриного зародыша на третьи сутки инкубации удалить скорлупу, поместить его в пластиковую оболочку и культивировать до конца эмбриогенеза, то в условиях дефицита кальция большая часть хрящевого скелета не превращается в костную ткань (Tuan. Lynch. 1983; рис. 6.13). Остеогенез в длинных костях начинается в диафизе и постепенно распространяется к концам хрящевого зачатка. У новорожденного младенца диафизы длинных костей обызвествлены, тогда как их концы (называемые эпифизами) все еще образованы хрящом. Вскоре после рождения в хрящевых эпифизах (по мере того как в них проникают кровеносные сосуды) возникают вторичные центры окостенения. Между кальцинированной тканью в эпифизах и диафизом продолжает сохраняться хрящевая пластинка. Эта эпифизарная пластинка играет важную роль в росте ребенка после рождения, так как она отвечает пролиферацией на поступающий в нее гормон роста (рис. 6.14). Гормоны также ответственны и за прекращение роста: высокие концентрации эстрогена или тестостерона вызывают слияние центров окостенения в эпифизах и диафизе, что приводит к утрате эпифизарной пластинки. Такое эпифизарное слияние происходит в юношеском возрасте, причем у девушек на два года раньше, чем у юношей. По мере увеличения костного матрикса на периферии кости в ее внутренней части образуются полости и формируется костный мозг. Эта деструкция костной ткани осуществляется остеокластами многоядерными клетками, поступающими в кость по кровеносным сосудам (Kahn, Simmons, 1975).
| ||||||||