Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с._______________ РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. МЕЗОДЕРМА И ЭНТОДЕРМА____________ ___________ 191
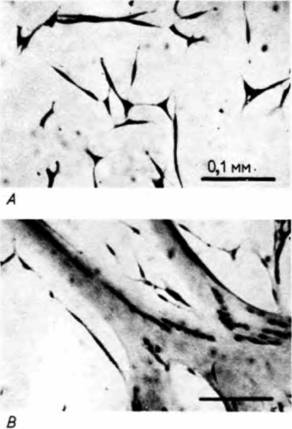
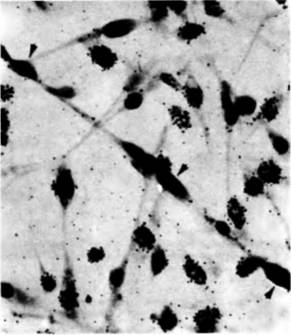
делить процесс узнавания (выстраивание миобластов в цепочку) от процесса слияния, обработав культивируемые клетки фосфолипазой С. Этот фермент препятствует слиянию мембран, но не действует на процесс узнавания. Поэтому в присутствии фермента клетки образовывали цепочки, но их мембраны не сливались. Эти опыты также показали, что процесс узнавания ответствен за остановку синтеза ДНК в миобласте. 3Н-тимидин включался в ДНК мигрирующих миобластов, но не включался в миобласты, расположенные в цепочку (рис. 6.7). Следовательно, процесс узнавания клетками приостанавливает их дальнейшее деление. Вторая стадия миогенеза это процесс слияния миобластов. Оба события — и узнавание, и слияние — зависят от наличия ионов кальция, но во время слияния ионы кальция должны поступать в миобласты. В соответствии с этим слияние можно активировать Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.192_______________ ГЛАВА 6_______________________________________________________________________________
ионофорами кальция, такими, как A23I87, который переносит ионы кальция через клеточные мембраны (Schainberg et al., 1969; David et al., 1981). Если выстраивание миобластов в цепочку опосредовано гликопротеидами клеточной мембраны (Knudsen, 1985), то их слияние, по-видимому, регулируется интенсивной реорганизацией мембранных компонентов, приводящей к образованию участков, свободных от белков и содержащих фосфолипиды (Kaufman, Foster, 1985; Wakelam, 1985). Было показано (Kalderon, Gilula, 1979), что опосредованное кальцием слияние миобластов происходит в тех участках их мембран, которые обогащены фосфолипидами. Эта гипотеза объясняет, почему фосфолипаза С является таким мощным ингибитором слияния клеток. Хотя узнавание клетками друг друга и их слияние — существенные черты миогенеза. видовая специфичность в этих явлениях отсутствует. Миобласты сливаются только с миобластами, и они не обязательно должны принадлежать одному и тому же виду животных. Показано (Yaffe, Feldman, 1965), что миобласты зародышей крысы и курицы легко сливаются, образуя гибридные мышечные трубочки. Результаты исследований, проведенных на аллофенных мышах, также свидетельствуют в пользу гипотезы о слиянии миобластов. Таких мышей получают путем слияния двух ранних зародышей; у развивающихся мышей имеются две четко различимые популяции клеток (см. рис. 3.29). В одном из опытов (Mintz, Baker, 1967) объединяли зародышей двух линий мышей, у которых синтезируются разные формы фермента изоцитратдегидрогеназы. Этот фермент, обнаруженный во всех клетках, состоит из двух идентичных субъединиц. Если мышечные трубочки образуются из одной клетки, ядра которой делятся без цитокинеза, то следует ожидать появления у аллофенной мыши двух разных («родительских») форм фермента (рис. 6.8). Вместе с тем, если мышечные трубочки формируются путем слияния миобластов, то следует ожидать появления мышечных клеток, в которых экспрессируются гены не только одной из двух «родительских» изоформ фермента (АА или ВВ), но также и третьей изоформы, образованной субъединицами каждого из родительских типов фермента (AB). Различные формы изоцитратдегидрогеназы могут быть разделены и идентифицированы по их подвижности в электрическом поле. Результаты опыта ясно показали наличие гибридного фермента в экстрактах из скелетных мышц, хотя в экстрактах из всех других тканей был обнаружен фермент только двух родительских типов. Следовательно, мышечные трубочки действительно формируются путем слияния многочисленных миобластов. Все исследователи считают слияние миобластов и синтез специфических мышечных белков независимыми друг от друга явлениями, хотя и происходящими обычно одновременно. Как только миобласты выстраиваются в цепочку и прекращают делиться. они начинают синтезировать тропомиозин, тяжелую цепь миозина и специфичные для мышц изоформы актина и креатинкиназы. Креатинкиназа представляет собой димерный белок. До слияния миобластов в них синтезируется креатинкиназа, состоящая только из субъединиц В (димер ВВ). К моменту слияния в культуре они начинают синтезировать другую специфичную для мыши форму фермента (М), создавая димеры ММ (рис. 6.9. А). Слияние миобластов можно стимулировать, заменив среду, поддерживающую пролиферацию, средой, в которой отсутствуют определенные факторы роста. В лимитированной среде миобласты прекращают делиться и в течение короткого периода времени сливаются. Эта методика была использована (Devlin, Emerson, 1978) для синхронизации слияния клеток и изучения синтеза миозина, актина и тропомиозина (рис. 6.9, Б). В начале культивирования ни одного из белков обнаружить не удается. Однако ко времени слияния миобластов эти структурные белки очень быстро выяв- |