|
Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
|
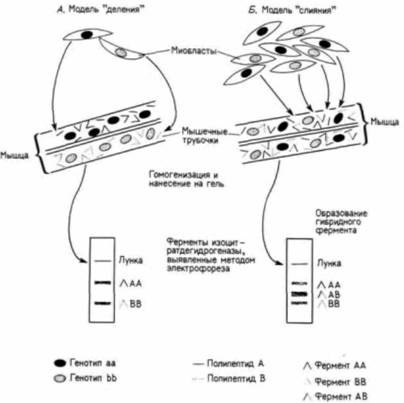
Рис. 6.8. Схема, иллюстрирующая два возможных механизма формирования скелетных мышц и способ выбора между ними. Аллофенных мышей получают путем слияния мышиных зародышей разных линий: клетки каждой из двух линий можно различить по наличию разных форм фермента изоцитратдегидрогеназы. Этот фермент состоит из двух субъединиц; одна линия мышей продуцирует А-субъединицы изоцитратдегидрогеназы, другая — В-субъединицы. А. Если ферменты синтезируются одной клеткой или в многоядерных клетках, образовавшихся путем деления ядер в одной клетке, то изоформы фермента будут либо АА, либо ВВ, Б. Если в одной и той же клетке имеются два разных ядра, то одно из них может кодировать субъединицу В, тогда как другое — субъединицу А, и в результате некоторые молекулы фермента будут гибридными (AB). Эти три типа молекул разделяют методом электрофореза. Присутствие молекулы AB только в клетках скелетных мышц (но не в других типах клеток) подтверждает правильность модели слияния. (По Mintz, Baker, 1967.)
|
|
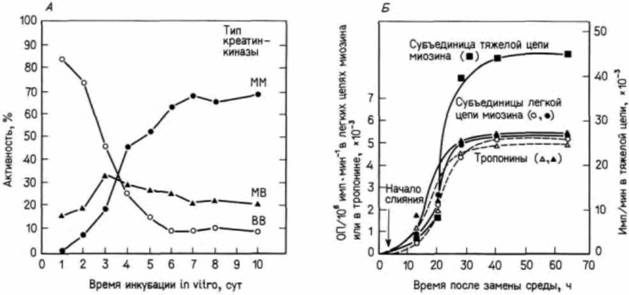
Рис. 6.9. Синтез новых ферментов, сопутствующий слиянию миобластов. А. Уменьшение активности субъединиц немышечной креатинкиназы (B-типа), сопровождающееся замещением их субъединицами специфичной для мышц креатинкиназы (М-типа) по мере слияния миобластов куриного зародыша, культивируемых in vitro. Слияние начинается после первых суток культивирования, и на вторые сутки культивирования в мышечных трубочках обнаруживается 38% ядер. Б. Синтез специфичных для мышц сократительных белков после перенесения делящихся миобластов в среду, способствующую их слиянию. Культуры метили 35S-метионином, который включается в белки, а затем через определенные сроки выявляли синтез разных белков (А — по Lough, Bischoff, 1977; Б —поDevlin, Emerson, 1978.)
|
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.
194_______________ ГЛАВА 6_______________________________________________________________________________
ляются. Показано (Sutherland, Königsberg, 1983), что, когда прекращается синтез ДНК, синтез специфичных для мышц белков увеличивается как обычно, даже в том случае, если слияние миобластов подавлено понижением концентрации кальция в среде. Отсюда можно заключить, что, как только клетки прекращают делиться, они приобретают способность и к слиянию, и к синтезу специфичных мышечных белков.
Общепризнано, что огромное увеличение синтеза таких белков происходит к моменту слияния миобластов, однако остается неясным, на каком уровне осуществляется контроль этих процессов — на транскрипционном, трансляционном или на том и другом вместе. Некоторые исследователи (см. гл. 14) сообщили, что в миобластах до их слияния обнаруживаются мРНК, специфичные для мышечных белков. Если это так, то акт слияния должен действовать как сигнал для начала трансляции этих уже имеющихся мРНК. Другие исследователи (Devlin, Emerson, 1979; Daubas et al., 1981 ) не смогли обнаружить в делящихся миобластах мРНК для сократительных белков. Эти исследователи полагают, что синтез таких белков происходит на новообразованных мРНК. В любом случае слияние миобластов и накопление специфичных мышечных белков, по-видимому, координируется прекращением синтеза ДНК в пролиферирующих миобластах.
Остеогенез: развитие кости
Кости наиболее четко выраженные производные соматической мезодермы. В этой главе будут рассмотрены только самые общие механизмы формирования костей. Студенты, желающие получить более подробные сведения, могут обратиться к учебникам по гистологии, где этой теме посвящены целые главы. Существуют два способа формирования костей (остеогенеза), и оба включают в себя трансформацию предсуществующей соединительной ткани в костную ткань. Превращение малодифференцированной соединительной ткани в костную называется интрамембранным окостенением, а замещение хряща костью — эндохондральным окостенением. Образование плоских костей, например костей черепа, осуществляется посредством интрамембранного окостенения. Рыхло расположенные мезенхимные клетки пролиферируют и образуют плотные скопления. Некоторые из этих клеток дают начало капиллярам, а другие изменяют свою форму и становятся остеобластами — клетками, способными секретировать костный матрикс. Синтезируемый ими коллагено-гликозаминогликановый матрикс обладает способностью связывать соли кальция, приносимые капиллярами в область окостенения. Таким образом, матрикс становится кальцифицированным (обызвествленным). В большинстве случаев остеобласты отделяются от области обызвествления слоем предкостного (остеоидного) матрикса. который они секретируют. Однако время от времени остеобласты захватываются костным матриксом, и тогда они становятся остеоцитами — костными клетками. По мере обызвествления костные спикулы распространяются радиально от центра, где началось окостенение (рис. 6.10). Кроме того, целая область обызвествленных спикул окружается плотно лежащими над ней мезенхимными клетками, формирующими периост, или надкостницу. Клетки на внутренней поверхности надкостницы также становятся остеобластами и секретируют костный матрикс параллельно с матриксом уже образовавшихся спикул. Таким образом формируется много слоев кости.
Эндохондральное окостенение заключается в замещении хрящевой ткани костной. В данном случае хрящ служит моделью для последующего формирования кости. Сначала образуется хрящевой зачаток
|
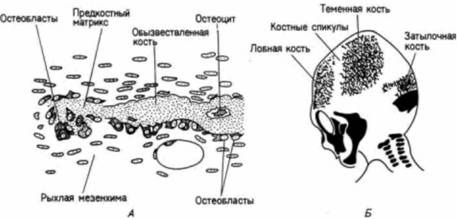
Рис. 6.10. Схема, иллюстрирующая внутримембранное окостенение. А. В скоплениях мезенхимы образуются остеобласты, которые секретируют костеподобный (предкостный) матрикс. Эти остеобласты располагаются вдоль обызвествленного участка матрикса. Остеобласты, попавшие внутрь костного матрикса, становятся остеоцитами. Б. Распространение костных спикул от места первичного окостенения в костях черепа трехмесячного зародыша человека. Кости, образующиеся путем эндохондрального окостенения, обозначены черным цветом. (По Langman, 1969.)
|
|
|
|
 Скачать 10.9 Mb.
Скачать 10.9 Mb.