БФ скелетных мышц. Биофизика мышечного сокращения
 Скачать 479.5 Kb. Скачать 479.5 Kb.
|

1 2 Государственное образовательное учреждение высшего профессионального образования Читинская Государственная Медицинская Академия федерального агентства по здравоохранению и социальному развитию Кафедра медицинской физики и информатики КУРСОВАЯ РАБОТА Тема: Биофизика мышечного сокращения Чита – 2009 г. ПЛАН I. Введение II. Сокращение скелетных мышц 1. Миография 2. Механические свойства мышцы 3. Изотоническое и изометрическое сокращения 4. Работа мышц 5. Методы раздражения мышц III. Потенциал покоя и потенциал действия скелетного мышечного волокна 1. Одиночное сокращение 2. Суммация сокращений и тетанус 3. Моторные единицы 4. Тонус скелетной мышцы IV. Механизм мышечного сокращения 1. Структура мышц 2. Роль АТФ в мышечном сокращении 3. Теории мышечного сокращения V. Работа и сила мышц VI. Утомление мышцы 1. Эргография VII. Гладкие мышцы 1. Особенности гладких мышц 2. Возбудимость и сокращение гладкой мышцы 3. Тонус гладкой мышцы 4. Автоматия гладких мышц VIII. Вывод I. Введение Вся жизнедеятельность животных и человека неразрывно связана с механическим движением, осуществляемым мышцами. Все телодвижения, кровообращение, дыхание и прочие акты возможны благодаря наличию в организме мышц, обладающих специальным белковым сократительным комплексом — актомиозином. Однако наличие сократительных элементов имеет значение не только при совершении вышеперечисленных макродвижений. В настоящее время накапливается все больше и больше данных о роли сократительных элементов в микропроцессах, в частности при активном транспорте веществ через мембраны и при движении цитоплазмы. Как было установлено, цитоплазма всех клеток находится в постоянном движении. По данным Камия, цитоплазма обладает колебательным, циркуляционным, фонтанирующим и другими видами движения, что, несомненно, играет большую роль в протекании метаболических процессов в клетках. В настоящее время нет единой точки зрения на причины происхождения этих движений цитоплазмы, однако наиболее вероятной является гипотеза функционирования сократительных элементов, подобных мышечным. II. Сокращение скелетных мышц мышца гладкая сокращение возбудимость Основными физиологическими свойствами мышц являются их возбудимость, проводимость и сократимость. Последняя проявляется или в укорочении мышцы, или развитии напряжения. Миография Для регистрации мышечного сокращения применяется методика миографии, т.е. графической регистрации сокращения с помощью рычажка, присоединенного к одному концу мышцы. Свободный конец рычажка чертит на ленте кимографа кривую сокращения – миограмму. Этот способ регистрации мышечного сокращения прост и не требует сложного оборудования, но имеет тот недостаток, что инерция рычажка и его трении по поверхности ленты кимографа несколько искажают запись. Во избежание этого недостатка теперь применяют специальный датчик, преобразующий механические изменения (линейные перемещения или усилия мышцы) в колебания силы электрического тока. Последние регистрируются с помощью шлейфного или катодного осциллографа. Точной методикой является также оптическая регистрация, производимая с помощью пучка света, отраженного от зеркальца, наклеенного на брюшко мышцы. По своим механическим свойствам мышцы относятся к эластомерам — материалам, обладающим эластичностью (растяжимостью и упругостью). Если мышцу подвергнуть действию внешней механической силы, то она растягивается. Величина растяжения мышцы в соответствии с законом Гука будет пропорциональна величине деформирующей силы (в определенных пределах): где Δl — абсолютное удлинение мышцы; l — начальная длина мышцы; F— деформирующая сила; S — площадь поперечного сечения мышцы; α — коэффициент упругости. Величина отношения F/Sназывается механическим напряжением, а величина l/α — модулем упругости; он показывает величину напряжения, необходимого для удлинения тела в 2 раза относительно начальной длины. По своим свойствам мышца приближается к каучуку, модуль упругости для обоих этих материалов равен примерно 10 кгс/см2. Мышцы обладают и другими свойствами, присущими каучуку. Как и при растяжении каучука, при сильном растяжении мышцы наблюдается локальная кристаллизация (упорядочение макромолекулярной белковой структуры фибриллярного типа). Это явление было изучено методом рентгеноструктурного анализа. При этом освобождается кристаллизационное тепло, в результате чего температура мышцы при растяжении повышается. После того как внешнюю силу убирают, мышца восстанавливает свою длину. Однако восстановление не бывает полным. Наличие остаточной деформации характеризует пластичность мышцы — способность сохранять форму после прекращения действия силы. Таким образом, мышца не является абсолютно упругим телом, а обладает вязкоупругими свойствами. При очень сильном растяжении мышца ведет себя как нормальное упругое тело. В этом случае при растяжении температура мышцы понижается. При сокращении мышцы развивается напряжение и совершается работа. Мышцы обладают сократительными и эластическими элементами. Поэтому возникающее напряжение и совершаемая работа обусловлены не только активным сокращением сократительного комплекса, но и пассивным сокращением, определяемым эластичностью или так называемым последовательным упругим компонентом мышцы. За счет последовательного упругого компонента работа совершается только в том случае, если мышца была предварительно растянута, и величина этой работы пропорциональна величине растяжения мышцы. Этим в большой степени объясняется то, что наиболее мощные движения совершаются при большой амплитуде, обеспечивающей предварительное растяжение мышц. Мышечные сокращения делятся на изометрические – происходящие при неизменной длине мышцы, и изотонические – происходящие при неизменном напряжении. Чисто изометрические или чисто изотонические сокращения с большим или меньшим приближением можно получить только в лабораторных условиях при работе на изолированных мышцах. В организме сокращения мышц никогда не бывают чисто изометрическими или чисто изотоническими. Скелетные мышцы с помощью сухожилий прикрепляются к костям, которые образуют систему рычагов. В большинстве случаев мышцы прикрепляются к костям так, что при их сокращении наблюдается выигрыш в амплитуде движений и эквивалентный проигрыш в силе. Плечо рычага мышцы в большинстве случаев бывает меньше плеча рычага соответствующей кости. Согласно Аккерману, механический выигрыш в амплитуде движений большинства конечностей человека имеет величину от 2,5 до 20. Для двуглавой мышцы плеча он равен приблизительно 10. При движении костей соотношение плеч рычагов мышц и костей меняется, что приводит к изменению напряжения мышц. По этой причине изотонических сокращений в естественных условиях не наблюдается. По этой же причине в процессе сокращения меняются вышеприведенные величины механического выигрыша в амплитуде движений. В зависимости от величины силы, которую преодолевает мышца, скорость сокращения (укорочения) мышцы бывает различной. Хилл на основе опытных данных, полученных при работе на изолированных мышцах, вывел так называемое основное уравнение сокращения мышцы. Согласно Хиллу, скорость сокращения мышцы vнаходится в гиперболической зависимости от величины нагрузки F: (F + a) (v + b) = const, где а и b — константы, приблизительно равные ¼ Fи соответственно ¼ v.  Нагрузка, кгс Рис.1. Зависимость скорости сокращения мышцы лягушки от величины нагрузки Байером были сделаны интересные замечания к уравнению. Уравнение приводится к виду F' v' = const, если принять F' = F + aи v' = v + b. Произведение F х v' представляет собой общую мощность, развиваемую мышцей при сокращении. Так как Fvменьше F'v', т. е. внешняя мощность меньше общей мощности, то следует предположить, что мышца совершает не только внешнюю работу, но еще и некоторую внутреннюю работу, проявляющуюся в том, что нагрузка как бы увеличивается на величину а, а скорость сокращения на величину b. Эту внутреннюю работу можно интерпретировать как потерю энергии на внутримолекулярное трение в форме теплового рассеивания. Тогда с учетом высказанных замечаний можно отметить, что общая мощность мышцы в физиологических пределах является постоянной величиной, не зависящей от величины нагрузки и скорости сокращения. С термодинамической точки зрения мышца представляет собой систему, которая преобразует химическую энергию (энергию АТФ) в механическую работу, т. е. мышца является хемо-механической машиной. Как уже отмечалось, при сокращении мышцы происходит теплообразование. Хиллом с помощью термоэлектрических методов было установлено, что при каждом раздражении вначале выделяется постоянная по величине и не зависящая от нагрузки теплота активации Q, а затем теплота сокращения kΔl, пропорциональная сокращению мышцыΔl и не зависящая от нагрузки (k-коэффициент пропорциональности). Если сокращение изотоническое, то мышца производит работу А, равную произведению нагрузки F на величину сокращения: А = FΔl. Согласно первому закону термодинамики, изменение внутренней энергии ΔU мышцы будет равняться сумме выделенного тепла и совершенной работы: -ΔU = Q + kΔl + FΔl = Q + Δl (F + k) Тогда КПД мышечного сокращения будет равен:  Учитывая, что величины Q и k не зависят от F, из последнего уравнения следует, что в определенных пределах КПД мышечного сокращения будет увеличиваться при увеличении нагрузки. Хилл на основе полученных им в опытах данных определил, что КПД мышечного сокращения примерно равен 40%. Если бы мышца работала как тепловая машина с КПД 40%, то при температуре среды 200С температура мышцы должна была бы быть равной 2150С. Величина КПД 40% показывает эффективность превращения энергии АТФ в механическую энергию. Если учесть, что КПД окислительного фосфорилирования, в процессе которого синтезируется АТФ, имеет величину около 50%, то полная эффективность превращения энергии питательных веществ в механическую энергию будет равна приблизительно 20%. Методы раздражения мышц. Для того чтобы вызвать сокращение мышцы, ее подвергают раздражению. Непосредственное раздражение самой мышцы (например, электрическим током) называется прямым раздражением; раздражение двигательного нерва, ведущее к сокращению иннервированной этим нервом мышцы, называется непрямым раздражением. Ввиду того, что возбудимость мышечной ткани меньше, чем нервной, приложение электродов раздражающего тока непосредственно к мышце еще не обеспечивает прямого раздражения: ток, распространяясь по мышечной ткани, действует в первую очередь на находящиеся в ней окончания двигательных нервов и возбуждает их, что ведет к сокращению мышцы. Чтобы получить сокращение мышцы под влиянием прямого раздражения, необходимо либо выключить в ней двигательные нервные окончания ядом кураре, либо прикладывать стимул через введенный внутрь мышечного волокна микроэлектрод. III. Потенциал покоя и потенциал действия скелетного мышечного волокна Мембранный потенциал покоя мышечного волокна составляет 80—90 мв, а потенциал действия—120—135 мв. Условием возникновения потенциала действия является критическая деполяризация мембраны. При непрямом раздражении мышцы критическая деполяризация мембраны осуществляется ацетилхолином, выделяющимся в нервных окончаниях при поступлении к ним нервного импульса. Возникший в области нервно-мышечного соединения потенциал действия распространяется далее вдоль всего мышечного волокна. При прямом раздражении электрическим током потенциал действия в мышечном волокне возникает под катодом и распространяется из этой точки по волокну. Длительность потенциала действия мышечного волокна — 3—5 мсек, т. е. в 5—10 раз больше продолжительности потенциала действия двигательных нервных волокон, иннервирующих мышцу. Поэтому максимальное число импульсов, которое может воспроизводить мышца, не превышает 200—250 в секунду. В возникновении потенциала покоя в скелетном мышечном волокне главную роль играют ионы К', диффундирующие из клетки в окружающую среду, и ионы Cl диффундирующие в противоположном направлении. Потенциал действия обусловлен повышением натриевой проницаемости мембраны и потоком ионов Na' внутрь волокна. Одиночное сокращение В ответ на одиночное раздражение — прямое или непрямое — мышца отвечает одиночным сокращением. Последнее подразделяют на три фазы: латентный период сокращения, фаза сокращения и фаза расслабления. Началу сокращения каждого мышечного волокна предшествует потенциал действия. 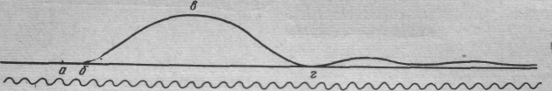 Рис. 2. Кривая одиночного сокращения скелетной мышцы а – момент раздражения; а - б – латентный период; б – в – период укорочения; в – г – период расслабления При обычном способе миографической регистрации сокращения мышцы латентный период составляет примерно 0,01 секунды. При более совершенных методах — при оптической регистрации сокращения — латентный период короче (около 0,0025 секунды). Поэтому некоторые исследователи полагают, что он равен или почти равен латентному периоду потенциала действия, т. е. что сокращение начинается одновременно или почти одновременно с потенциалом действия. Причина запаздывания записи сокращения по сравнению с действительным его началом заключается в том, что мышца начинает сокращаться не вся сразу: сначала мышца сокращается в месте раздражения, а при непрямом раздражении — в тех пунктах, где в ней расположены двигательные нервные окончания. Сократившаяся часть мышцы растягивает соседние участки, которые в силу своей растяжимости поддаются этой тяге. Таким образом, общая длина мышцы некоторое время не изменяется и рычажок миографа в первый момент остается неподвижным. Сокращение, так же как и возбуждение, распространяется вдоль мышечных волокон. В этом можно убедиться посредством следующего опыта: на мышцу лягушки с параллельными волокнами, например портняжную мышцу, помещают два рычажка и наносят раздражение на один конец мышцы. При прохождении волны сокращения рычажки приподнимаются по очереди: сначала ближайший к месту раздражения, затем дальний. Скорость проведения волн возбуждения и сокращения одинакова. Электрофизиологическим методом обнаружена скорость проведения возбуждения в двуглавой мышце плеча человека в пределах 3,5—5 м/сек. Ввиду относительно большой продолжительности сокращения наступает момент, когда вся мышца сокращена, так как последние ее участки, до которых доходит волна возбуждения, уже сократились, а первые еще не расслабились. Этот момент соответствует моменту максимального укорочения мышцы. Скорость сокращения разных мышц тела неодинакова. Так, у теплокровных при изометрическом режиме работы длительность одиночного сокращения глазодвигательных мышц составляет примерно 10 мсек, длительность сокращения икроножной мышцы — 50 мсек, а камбаловидной мышцы — 100 мсек. При изотоническом режиме работы мышцы скорость ее сокращения снижается при увеличении поднимаемого ею груза. Величина одиночного сокращения скелетной мышцы зависит от силы раздражения. При пороговом раздражении сокращение еле заметно, с увеличением же силы раздражения оно нарастает (субмаксимальное сокращение), пока не достигнет известной высоты, после чего высота остается неизменной, несмотря на увеличение силы раздражения (максимальное сокращение). Это объясняется тем, что мышца состоит из большого числа волокон. Каждое из мышечных волокон реагирует на раздражение по закону «все или ничего». Но так как отдельные волокна обладают неодинаковой возбудимостью, то только часть их возбуждается при слабом раздражении. Чем сильнее раздражитель, тем большее число волокон возбуждается. При максимальном сокращении сокращаются все волокна мышцы. Суммация сокращений и тетанус В естественных условиях в организме скелетная мышца получает обычно из нервной системы не одиночные раздражения, а ряд быстро следующих друг за другом нервных импульсов. Под влиянием ритмических сражений наступает сильное и длительное укорочение мышцы. Такое сокращение называется тетаническим сокращением, или тетанусом. То, что длительное укорочение действительно возникает в результате большого числа отдельных вспышек возбуждения, легко доказать, регистрируя потенциалы действия в тетанически сокращенной мышце. Так, при произвольных движениях конечностей человека число потенциалов действия, возникающих в его мышцах во время сокращения, составляет 50—70 в секунду. Тетанические сокращения мышцы представляют собой результат суммации одиночных сокращений. Для исследования суммации сокращений наносят на мышцу два одиночных раздражения. Промежуток времени между раздражениями должен быть такой, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого сокращения. При этом возможны два случая. Если второе раздражение поступает, когда мышца уже начала расслабляться после первого сокращения, то на миографической вершина второго сокращения будет отделена от вершины первого небольшим западением кривой. Если же второе раздражение действует, когда первое сокращение еще не дошло до своей вершины, то второе сокращение полностью сливается с первым, образуя вместе с ним единую суммированную вершину. Таким образом, возможны два типа суммации. Гладкий и зубчатый тетанус. Для искусственного воспроизведения тетануса на мышцу действуют большим числом раздражений, следующих друг за другом с такой частотой, при которой происходит суммация. При относительно малой частоте наступает зубчатый тетанус, при большой частоте – гладкий тетанус. После прекращения тетанического раздражения волокна вначале не полностью расслабляются, и их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется послететанической, или остаточной, контрактурой. Моторные единицы Каждое моторное нервное волокно, являющееся отростком двигательной клетки передних рогов спинного мозга, иннервирует не одно, а целую группу мышечных волокон. Такая группа получила название моторной единицы. Количество мышечных волокон, входящих в состав моторной единицы в разных мышцах человека, варьирует от 10 до 3000. Наименьшее число волокон содержится в моторных единицах быстрых мышц, обеспечивающих наиболее точные движения. Так, в глазных мышцах и мышцах пальцев руки моторные единицы имеют в своем составе 10—25 мышечных волокон, причем каждое из них получает иннервацию со стороны нескольких нервных волокон. В отличие от этого относительно медленные мышцы, участвующие в регуляции позы тела и ненуждающиеся в точном контроле, состоят из моторных единиц, включающих в свой состав от 2000 до 3000 волокон. Моторные единицы икроножной мышцы содержат около 1500 волокон. Вследствие того что скорость распространения возбуждения в нервных волокнах, иннервирующих скелетные мышцы, очень велика, мышечные волокна, составляющие моторную единицу, приходят в состояние возбуждения практически одновременно. Электрическая активность моторной единицы имеет вид частокола, в котором каждому пику соответствует суммарный потенциал действия многих одновременно возбужденных волокон. Для того, чтобы зарегистрировать электрическую активность моторной единицы, необходимо вколоть в брюшко мышцы очень тонкие электроды, которые на всем своем протяжении, за исключением самого кончика, покрыты изолирующим лаком. С помощью таких электродов регистрируют активность одной моторной единицы различных мышц человека. Суммация сокращений моторных единиц в целой мышце. В отличие от мышечных волокон каждой моторной единицы, синхронно, т. е. одновременно, возбуждающихся в ответ на приходящий импульс, мышечные волокна различных моторных единиц мышцы, как правило, работают асинхронно. Объясняется это тем, что моторные единицы иннервируются различными двигательными нейронами, которые посылают импульсы с разной частотой и разновременно. Несмотря на неодновременность начала и конца сокращения различных моторных единиц, суммарное сокращение мышцы в целом имеет в условиях нормальной деятельности слитный характер, по форме своей напоминающий гладкий тетанус, даже в том случае, когда каждая из моторных единиц работает в редком ритме (например, 5 сокращений в секунду). Таким образом, при асинхронной деятельности моторных единиц, обусловленной асинхронной работой соответствующих нейронов спинного мозга, все движения нашего тела имеют плавный характер уже при малой частоте двигательной импульсации. Асинхронная деятельность моторных единиц не позволяет различать электрическую активность каждой из них при отведении потенциалов от целой мышцы: неодновременно возникающие пики потенциалов действия алгебраически суммируются (интерферируют) на электродах, вследствие чего возникает сложная картина, по которой можно лишь косвенно судить о степени возбуждения отдельных мышечных волокон. В покое моторные единицы мышц конечностей человека обнаруживают лишь очень редкие разряды потенциалов действия. Это обусловливает тонус мышц. При небольшом напряжении появляются разряды с частотой 5—10 в секунду. Повышение напряжения увеличивает частоту следования потенциалов действия до 20—50 в секунду. Сила мышечного сокращения зависит от числа моторных единиц, вовлекаемых одновременно в реакцию, и от частоты возбуждения каждой из них. Тонус скелетных мышц В покое, вне работы, мышцы не являются полностью расслабленными, а сохраняют некоторое напряжение, называемое тонусом. Внешним выражением тонуса является определенная степень упругости мышц. Электрофизиологические исследования показывают, что тонус скелетных мышц связан с поступлением к мышце отдельных следующих друг за другом с большим интервалом нервных импульсов, возбуждающих попеременно различные мышечные волокна. Эти импульсы возникают в мотонейронах спинного мозга, активность которых в свою очередь поддерживается импульсами, исходящими как из вышележащих центров, так и с периферии от рецепторов растяжения («мышечных веретен»), находящихся в самих мышцах. О рефлекторной природе тонуса скелетных мышц свидетельствует тот факт, что перерезка задних корешков, по которым чувствительные импульсы от мышечных веретен поступают в спинной мозг, приводит к полному расслаблению мышцы. У человека тонус мышц в известных пределах может регулироваться произвольно - по желанию человек, может почти полностью расслабить мышцы или же несколько напрячь их, не совершая, однако, при этом движения. Имеется много данных, свидетельствующих, что в осуществлении тонуса скелетных мышц особую роль играют так называемые медленные— тонические волокна. От тетанических волокон они отличаются малой скоростью процессов сокращения и расслабления и большой способностью к суперпозиции ответов. Вследствие этого даже редкого ритма раздражений оказывается достаточным для длительного удержания мышечного волокна к укороченном состоянии. IV. Механизм мышечного сокращения Тонкая структура мышц. Скелетная мышца позвоночных состоит из нескольких тысяч параллельных мышечных волокон диаметром 10—100 мкм, заключенных в общую оболочку. К каждому мышечному волокну через концевую пластинку присоединено окончание нервного волокна. Мышечное волокно способно к сокращению под действием нервного импульса и представляет собой функциональный элемент мышечной системы. Протяженность волокна может быть равна длине самой мышцы или значительной ее части. Волокна на каждом конце мышцы переходят н сухожилие, которое принимает на себя напряжение при сокращении. Мышечное волокно в свою очередь содержит 1000—2000 параллельных мышечных фибрилл (миофибрилл) диаметром около 1 мкм. Весь пучок миофиорилл обтянут мембраной мышечного волокна — плазмалеммой. Плазмалемма, подобно мембранам всех других клеток, состоит из трех слоев белков и липидов общей толщиной около 10 нм и электрически поляризована. Мембранный потенциал достигает 100 мВ. Сверху плазмалемма покрыта тонким слоем коллагеновых нитей, обладающих упругими свойствами. В мышечном волокне содержится много ядер, располагающихся вблизи плазмалеммы, и большое количество митохондрий, находящихся между фибриллами. Митохондрии являются центрами образования макроэргических соединений, прежде всего АТФ. Отсюда макроэргические соединения через саркоплазму поступают к фибриллам. При микроскопическом исследовании видно, что в скелетных мышечных волокнах правильно чередуются темные и светлые полосы, образуя характерную поперечную полосатость. Поперечная полосатость волокон обусловлена поперечной полосатостью миофибрилл, расположенных строго определенно друг подле друга. Применяя метод электронного микроскопировапия и метод рентгеноструктурного анализа, удалось выяснить, что каждая миофибрилла состоит из параллельно лежащих толстых и тонких нитей — протофибрилл, чередующихся строго определенным образом. Дальнейшие исследования позволили установить, что толстые нити образованы молекулами белка миозина, а тонкие молекулами белка актина. Длина миозиновых нитей составляет примерно 1,5 мкм, а актиновых 1 мкм; толщина – соответственно 16 и 5-7 нм. В результате чередования толстых и тонких нитей возникает поперечная исчерченность, видимая под микроскопом. Для микроскопической картины поперечнополосатой мышцы характерно чередование плотных анизотропных полос (их называют А-диски) и светлых изотропных полос (I-диски). В А-дисках миозиновые нити образуют гексагональную (шестиугольную) упаковку; именно они обусловливают высокую оптическую плотность дисков. Активные нити прикрепляются с каждой стороны к узкой белковой структуре, так называемой Z-мембране, которая пересекает I-диск. Отрезок миофибрилл, заключенный между двумя Z-мембранами, называется саркомером. В мышечном волокне в том месте, где оба типа протофибрилл накладываются друг на друга, тонких протофибрилл в пучке в 2 раза больше, чем толстых. Тонкие протофибриллы оканчиваются у края Н-зоны – области с более низкой оптической плотностью, находящейся в середине А-диска. В центре А-диска расположена узкая темная полоска, известная под названием линии М. Считают, что эта линия соответствует небольшому утолщению, которое имеется в центре каждой толстой нити. Как показали Хэнсон и Леви, актиновые протофибриллы имеют форму двойной спирали, образованную глобулярными молекулами актина. Вся структура напоминает две плотные нитки бус, закрученные одна вокруг другой, где роль одной бусинки играет глобулярная молекула актина. Миозиновые протофибриллы также представляют собой результат агрегации отдельных молекул миозина. До настоящего времени окончательно не выяснено, как происходит соединение молекул миозина в протофибрилле. При увеличении до 600000 раз на микрофотографиях продольного среза мышцы можно видеть, что пары толстых и тонких протофибрилл соединены поперечными мостиками. Эти поперечные мостики являются единственным связующим звеном между протофибриллами и обеспечивают структурную целостность мышцы. В дальнейшем в результате применения метода рентгеноструктурного анализа было показано, что мостики образованы отростками миозиновых нитей, расположенных с интервалом 6—7 нм. Мостики соединяют толстую нить с каждой из шести тонких нитей, располагаясь по спирали, витки которой повторяются через каждые 40 нм. В центральной части миозиновых протофибрилл мостики отсутствуют и на электронной микрофотографии этим участкам соответствует «псевдо Н-зона», обладающая более низкой оптической плотностью, чем Н-зона. Ферментативные свойства актомиозина. Кальциевый насос. В.А. Энгельгардтом и М.Н. Любимовой (1939) было сделано очень важное открытие; они показали, что наряду с сократительными свойствами миозин обладает ферментативными свойствами, являясь ферментом аденозинтрифосфатазой, расщепляющей АТФ. В миофибриллах через поперечные мостики миозин образует комплексное соединение с актином. Энергия, выделяющаяся в процессе гидролиза АТФ, непосредственно используется для сокращения актомиозинового комплекса. Ферментативная активность актомиозина примерно в 10 раз выше активности одного миозина. Ферментативная активность, а следовательно, и способность к сокращению актомиозинового комплекса сильно зависят от присутствия в среде ионов кальция. Многие ученые считают, что в отсутствие ионов кальция актомиозин вообще не способен расщеплять АТФ и сокращаться. При увеличении концентрации кальция до определенного предела активность актомиозина увеличивается и достигает максимального значения при концентрации кальция, равной концентрации АТФ в среде. Предполагают, что ионы кальция входят в состав активных центров миозина, локализованных в области поперечных мостиков, и только после этого миозин проявляет АТФ-азную активность. Непосредственной причиной, вызывающей расщепление АТФ и сокращение миофибрилл, служит появление свободных ионов кальция в саркоплазме. Так, инъекция раствора, содержащего ионы кальция, в саркоплазму приводит к сокращению мышечного волокна при отсутствии нервного импульса и потенциала действия мышечного волокна. Наконец, с помощью специальных индикаторов кальция было показано, что в момент сокращения волокна происходит увеличение концентрации ионов кальция в саркоплазме. Согласно современным представлениям, в клетках функционирует специальный кальциевый насос, работа которого вызывает сокращение и расслабление миофибрилл. Этот насос, по мнению Бендолла, локализован в мембранах саркоплазматического ретикулума (эндоплазматической сети) мышечного волокна. Саркоплазматический ретикулум состоит из поперечно и продольно расположенных в саркоплазме трубочек, цистерн, пузырьков, стенки которых имеют типичное мембранное строение. Поперечная система саркоплазматического ретикулума представляет собой впячивание плазмалеммы, идущие внутрь в виде трубочек и охватывающие каждую фибриллу на уровне соединения А- и I-дисков в мышцах млекопитающих и на уровне Z-мембран у холоднокровных. По поперечным трубочкам саркоплазматического ретикулума возбуждение в виде волны деполяризации передается от поверхности волокна, возбуждаемой нервным импульсом, к миофибриллам. Это подтверждается классическим опытом Хаксли с локальным раздражением мышечного волокна лягушки. Микроэлектродом наносили очень слабое подпороговое раздражение на различные участки волокна. Локальное сокращение нескольких миофибрилл возникало только в случае нанесения раздражения на уровне Z-мембран, где локализованы трубочки поперечного саркоплазматического ретикулума. От поперечного ретикулума возбуждение передается расположенному между фибриллами продольному ретикулуму, где локализован кальциевый насос. Предполагается, что в процессе проведения возбуждения по мембранам ретикулума основную роль играют не ионы натрия и калия, а ионы кальция и магния. Деполяризация мембран трубочек и пузырьков саркоплазматического ретикулума приводит к освобождению содержащихся в них моном кальция. Механизм освобождения ионов кальция пока не установлен. Возможно, это связано с увеличением проницаемости мембран для ионов кальция при возбуждении и последующей диффузией их по концентрационному градиенту в саркоплазму. Появление свободных ионов кальция в саркоплазме приводит к проявлению АТФ-азной активности актомиозина и к сокращению миофибрилл. Для сокращения миофибрилл необходимо также наличие ионов магния, механизм действия которых пока не установлен. Процесс расслабления миофибрилл связан с удалением ионов кальция из саркоплазмы, осуществляемым саркоплазматическим ретикулумом. Элементы ретикулума обладают способностью к активному поглощению ионов кальция из окружающего раствора. Препараты саркоплазматического ретикулума, выделенного из мышц путем дифференцированного центрифугирования их гомогенатов, обладают способностью поглощать ионы кальция из раствора. При этом в некоторых случаях концентрация кальция внутри пузырьков и цистерн ретикулума превышала концентрацию кальция в окружающем растворе в 2000 раз. Наличие активного переноса кальция при расслаблении миофибрилл подтверждается и тем, что концентрация кальция в саркоплазме после микроинъекции начинает постепенно уменьшаться, что сопровождается расслаблением миофибрилл. Возможно, как предполагает Бендолл, что обратный перенос кальция связан с самим движением протофибрилл при сокращении, что исключает необходимость наличия специального механизма активного переноса кальция. Прежнее представление, согласно которому расслабление вызывается освобождением специфического фактора расслабления — фактора Марша, оказалось ошибочным. Этот фактор выделялся путем экстракции из гомогенатов мышц. Он содержал ферменты, имеющиеся и саркоплазме, и фрагменты ретикулума. Один из этих ферментов и был принят за фактор расслабления, хотя на самом деле расслабляющее действие оказывали фрагменты ретикулума. Необходимо отметить, что расслабление миофибрилл при удалении ионов кальция из саркоплазмы происходи только в том случае, если в саркоплазме содержится АТФ. Удаление АТФ из саркоплазмы приводит к возникновению между актином и миозином сильных электростатических связей, что обусловливает окоченение (контрактуру) мышцы и потерю ею способности к растяжению. Таким образом, сокращение миофибрилл вызывается расщеплением АТФ в присутствии ионов кальция, а расслабление – поступлением новых молекул АТФ к протофибриллам при отсутствии ионов кальция. Регулятором сокращения и расслабления миофибрилл является поступление ионов кальция в саркоплазму и их удаление в саркоплазматический ретикулум. Восстановление первоначальной длины мышцы после сокращения обусловлено, вероятно, наличием упругих элементов в мышечных волокнах и работой мышц антагонистов. Упругими элементами мышечного волокна являются коллагеновая оболочка, покрывающая плазмалемму, и, возможно, саркоплазматический ретикулум. Если с волокна снять сарколемму и заставить его сократиться, то волокно не может расслабиться спонтанно, хотя легко вытягивается до первоначальной длины при действии внешней силы. Теории механизма мышечного сокращения. До получения данных о тонкой структуре мышц процессы мышечного сокращения пытались объяснить деформацией изолированных молекулярных цепей белков, т. е. удлинением или укорочением отдельных белковых молекул или агрегатов молекул. Часто данные о деформации различных полимеров переносили на мышечное сокращение, без учета структуры мышечных волокон. Известно много полиэлектролитных полимерных систем, обладающих способностью к изменению длины при изменении химического состава окружающего раствора. Примером такой системы является вытянутая цепочка полиакриловой кислоты. При подкислении раствора такая цепочки сокращается, в щелочной среде она, наоборот, растягивается. Если подвесить к ней груз, то можно получить машину, совершающую механическую работу при изменении рН раствора. Существуют также редокс-модели и ионные модели мышц, в которых факторами сокращения являются соответственно изменения редокс-потенциала и концентрации свободных ионов. Во всех этих моделях изменение длины полимеров происходит в основном в результате изменения электростатического взаимодействия между звеньями полимера или между витками спирали и случае спиральных структур. Существует множество гипотез, пытающихся объяснить мышечное сокращение на основе свойств индивидуальных молекулярных цепей сократительных белков. Все эти гипотезы исходят из представления, что в основе сокращения мышцы лежат процессы конформационных изменений структуры белковых цепей. Так, Мейер еще в 1929 г. выдвинул гипотезу, согласно которой мышечное сокращение обусловлено деформацией пептидных цепей вследствие изменения электростатического взаимодействия ионогенных групп СООН и NH2 при изменении рН. В настоящее время считают, что изменение рН при возбуждении миофибрилл недостаточно, чтобы вызвать конформационные переходы белков, по может быть достаточно для освобождения ионов кальция, которые уже могут вызвать деформацию белковой цепи. Согласно другому представлению, акт сокращения представляет собой конформационный переход белковой структуры от α-конфигурации, когда нити линейно вытянуты, к β-конфигурации, когда нити собраны в клубок. Однако эти гипотезы не смогли объяснить реальную картину сложного строения мышечного волокна на молекулярном уровне, полученную в последнее время. Возможно, что при медленном сокращении гладких мышц происходит фактическая деформация (активное сокращение отдельных протофибрилл) белковых цепей, как считает Г.М.Франк, однако для сокращения скелетных мышц гораздо более обоснованными являются представления, исходящие из гипотезы скольжения нитей. Г.Хаксли и Хэнсон выдвинули гипотезу скольжения нитей. Ими было отмечено, что в широком интервале деформаций как при сокращении, так и при растяжении миофибрилл ширина А-диска остается постоянной. Напротив, при изменении длины саркомера изменяется ширина I-диска. Светлая Н–зона в А-диске также изменяется, но замечательно, что до тех пор, пока она существует, расстояние от конца одной Н-зоны через Z-мембрану до начала следующей Н-зоны (а это расстояние равно длине тонких нитей в миофибрилле) также остается постоянным. Если вспомнить, что А-диски образованы нитями миозина, а тонкие нити состоят их актина, то можно заключить, что в большой области деформаций мышцы длина миозиновых и актиновых нитей остается постоянной. Это можно объяснить только тем, что при сокращении мышцы нити просто скользят друг относительно друга без изменения своей длины. При сильном сокращении мышцы в середине А-диска появляется плотная зона, причем ширина этой зоны увеличивается по мере сокращения мышцы. Эта плотная зона появляется после полного исчезновения Н-зоны. Уменьшение Н-зоны при сокращении вызывается скольжением тонких нитей навстречу друг другу к центру А-диска. Измерив расстояние от Z-мембраны до противолежащего конца ноной плотной зоны (полосы сокращения), Г. Хаксли и Хчпсоп обнаружили, что оно равно половине длины тонкой протофибриллы. На этом основании они предположили, что новая зона соответствует тому участку саркомера, где концы противолежащих тонких нитей перекрываются друг с другом. Это предположение подтвердилось тем, что на микрофотографии поперечного среза мышцы в области новой плотной зоны было обнаружено в 2 раза больше тонких нитей, чем в остальной области А-диска. Кроме того, при сильном сокращении мышцы, после исчезновения I-диска в области Z-мембран также появляются темные полосы. Это объясняется тем, что миозиновые нити достигают Z-мембран и после этого происходит их деформация. В дальнейшем данные электронного микроскопирования были подтверждены результатами рентгеноструктурного анализа. Основные рефлексы рентгенограммы не изменяются при сокращении мышц. Это указывает на то, что длина нитей при сокращении не меняется. Приведенные данные очень важны, так как в отличие от электронно-микроскопических исследований, проводимых на фиксированных препаратах мышц, рентгенографические исследования проводились и на живых сокращающихся мышцах, и на нефиксированных ее препаратах. Перемещение тонких нитей относительно толстых происходит, при помощи мостиков, соединяющих миозиновые нити с актиновыми. Так как изменений в длине толстых и топких нитей во время сокращения не происходит, то из модели скольжения нитей вытекает, что конформационные изменения, порождающие движение, должны происходить в указанных мостиках, связывающих толстые и тонкие нити. Весь процесс сокращения имеет циклический характер. Миозиновые мостики прикрепляются к активным участкам актиновых нитей и под действием энергии гидролиза АТФ укорачиваются или изменяют угол наклона к миозиновым нитям, что приводит к определенному перемещению нитей друг относительно друга. Затем происходит отсоединение мостиков в данных участках актиновых нитей и присоединение их в новых участках. Этот циклический процесс повторяется многократно, в результате чего происходит непрерывное перемещение нитей друг относительно друга. Рентгенографические исследования подтвердили предположение о движении мостиков. По мнению Г.Хаксли, расщепление одной молекулы АТФ приводит к одному замыканию и размыканию мостиков и к перемещению нитей на один элементарный участок. 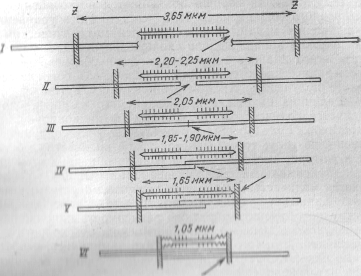 Рис.3. Схема, показывающая степень перекрывания нитей миозина и актина в поперечнополосатом мышечном волокне при различной длине саркомера Величина напряжения, развиваемого мышцей, определяется количеством замыкаемых (функционирующих) мостиков. Если мышца преодолевает при сокращении внешнюю силу, то замыкается такое количество мостиков, которое необходимо для уравновешивания этой силы. Максимальная сила, развиваемая мышцей, определяется количеством мостиков, которые могут замыкаться в данных условиях. Исходя из этих представлений, нетрудно объяснить обратную зависимость напряжения, развиваемого мышцей при сокращении, от скорости сокращения. Для того чтобы мостики замкнулись, необходимо какое-то время. При увеличении скорости скольжения нитей количество замыкаемых мостиков уменьшается, что обусловливает уменьшение напряжения, развиваемого мышцей. В зависимости от длины саркомеров длина участков, в которых нити актина и миозина перекрываются друг с другом, будет различной и, следовательно, будет различно количество мостиков, участвующих и создании напряжения, развиваемого мышцей. Учитывая, что максимальная сила миофибриллы определяется количеством функционирующих мостиков, следует ожидать, что максимальная сила изометрического сокращения миофибриллы будет изменяться с изменением длины саркомера. При длине саркомера 3,65 мкм нити актина и миозина уже не накладываются друг на друга и можно ожидать, что волокно не будет способно развивать силу. Под силой сокращения следует понимать разность между общей силой, развиваемой при раздражении мышцей, и упругой восстанавливающей силой, обусловленной эластическими элементами мышцы в случае се растяжения сверх нормальной длины. По мере сближения Z-мембран нити актина все глубже проникают в промежутки между нитями миозина и, наконец, при расстоянии 2,2 мкм все мостики миозиновых нитей приходят в контакт с нитью актина. Если именно эти мостики ответственны за возникновение силы, то следует ожидать, что в диапазоне от положения I до положения II сила будет пропорциональна степени перекрывания нитей. При дальнейшем укорочении волокна число мостиков, которые могут замыкаться, не изменяется и сила должна оставаться постоянной, пока длина саркомера не уменьшится до 2,05 мкм. В этот момент нити актина сходятся своими концами и сила должна убывать вследствие того, что тонкие нити, которые проникли дальше середины А-диска, будут неправильно ориентированы по отношению к миозиновым мостикам. Сила должна постепенно убывать, пока расстояние не достигнет 1,65 мкм, когда концы миозиновых нитей приходят в соприкосновение с Z-мембранами. При дальнейшем сокращении нити миозина должны деформироваться; сила должна убывать быстрее и совсем исчезать, когда актиновые нити доходят до противолежащих Z-мембран. Все эти предположения подтвердились экспериментально. Гордоном, А.Хаксли, Юлианом (1966) измерялось напряжение, развиваемое мышечным волокном при изометрическом сокращении, и одновременно методом фазово-контрастной микроскопии регистрировалась длина саркомера.  Рис.4. Зависимость силы сокращения поперечнополосатого мышечного волокна от длины саркомера Однако, несмотря на большие успехи в изучении механизма мышечного сокращения, все еще окончательно не установлен механизм работы мостиков, в результате которой энергия гидролиза АТФ превращается в механическую работу. В настоящее время имеется ряд гипотез, пытающихся объяснить конкретный механизм взаимодействия актиновых и миозиновых нитей. Наиболее глубоко разработанной и обоснованной является гипотеза Дэвиса. Согласно этой гипотезе, мостик между миозиновой и актиновой нитями образован полипептидными цепочками конца миозиновой молекулы, скрученными в спираль. В покое мостик вытянут—спираль находится в растянутом состоянии. Это обусловлена электростатическим отталкиванием двух отрицательных зарядов. Один из них находится в фиксированном состоянии у основания мостика, которое обладает АТФ-азной активностью. Другой отрицательный заряд локализован па конце мостика, с которым связана молекула АТФ. При возбуждении мышцы саркоплазматический ретикулум освобождает ионы кальция. Они образуют связь между молекулой АТФ, находящейся на конце мостика, и молекулой АДФ, расположенной на актиновой нити, что вызывает нейтрализацию отрицательных зарядов. Электростатическое отталкивание исчезает и растянутая цепочка — мостик — скручивается в α-спираль благодаря образованию водородных связей. Этот процесс представляет собой освобождение потенциальной энергии, запасенной вытянутой полипептидной цепочкой при первоначальном отталкивании зарядов. Укорочение полипептидной цени с образованием α-спирали приводит к двум эффектам. Во-первых, актиновая нить перемещается относительно миозиновой на один шаг; во-вторых, присоединенная молекула АТФ перемещается в область гипотетического АТФ-азного центра. Благодаря соответствующему расположению этого центра и наклону мостиков относительно толстой нити актиновые нити перемещаются в сторону М-линий. После этого АТФ расщепляется на АДФ и минеральный фосфат, что ведет к разрыву связей между актином и миозином. На место молекулы АДФ в миозиновом мостике из саркоплазмы поступает новая молекула АТФ, которая отталкивается отрицательным фиксированным зарядом миозина. В результате этого α-спираль растягивается – мостик удлиняется. Если в саркоплазме в это время имеются свободны ионы кальция, то весь цикл повторяется сначала. При этом во взаимодействии участвует уже следующий участок активной нити. Если же ионы кальция к этому времени удалены из саркоплазм, то волокно расслабляется. Модель Дэвиса получила ряд дополнений и подверглась модификациям. Бендолл (1970) предполагает, что присоединение ионов кальция в области мостиков приводит к изменению электрического взаимодействия. Нейтрализация отрицательных зарядов и присоединение актина к миозину обусловливают превращение спирали полипептидной цепочки (мостика) молекулы миозина в более беспорядочную, сильно свернутую конформацию но типу перехода «спираль — клубок». Такой переход сопровождаемся освобождением потенциальной (свободной) энергии, запасенном и более упорядоченной структуре — спирали. Эта энергия частично расходуется на тянущее усилие— перемещение нити актина на один шаг, а частично деградирует в тепло. Изменение конформации мостика одновременно вызывает сближение АТФ с АТФ-азным участком миозина, что вызывает гидролиз АТФ. 1 2 |