|
|
коллоквиум по биологии. колок. Человек как объект биологии. Значение биологического и социального наследования человека для медицины
Начиная с конца 80-х годов XX века получены убедительные доказательства связи некоторых видов наследственной патологии у человека с мутациями митохондриальной ДНК (см. гл. 4.1) В зависимости от типа мутаций митохондриальные болезни разделяют на 4 группы:
а) болезни, вызванные точковыми мутациями, приводящими к замене консервативных аминокислот в собственных белках митохондрий. К ним относятся пигментный ретинит и нейроофтальмопатия Лебера, при которой наступает двусторонняя потеря зрения. Выраженность клинических признаков у больных этими заболеваниями коррелирует с количеством мутантной мтДНК, которое у разных больных может варьировать от 5 до 100% всей мтДНК;
б) болезни, вызванные мутациями в генах т-РНК, приводящими к многочисленным дегенеративным заболеваниям с различной степенью тяжести клинических проявлений, коррелирующей с количеством мутантной мтДНК;
в) болезни вызванные делениями и дупликациями участков митохондриалъных генов. У человека описано тяжелое заболевание молодого и среднего возраста — отсроченная кардиопатия, при которой обнаружены делеции мтДНК кардиоцитов. Заболевание носит семейный характер. В ряде случаев предполагается Х-сцепленное наследование, что позволяет думать о существовании ядерного гена, мутация которого вызывает делению до 50% мтДНК кардиоцитов
г) болезни, вызванные снижением числа копий мтДНК, что является следствием определенных мутаций. К данной группе относятся летальная инфантильная дыхательная недостаточность и синдром молочнокислого ацидоза, при которых число копий мтДНК снижается до 1—2% от нормы. Снижение содержания мтДНК в клетках различных органов приводит к развитию миопатий, нефропатий, печеночной недостаточности и т.д. вследствие ослабления синтеза белков, кодируемых мтДНК.
Изменения в ДНК митохондрий сопровождаются нарушением их функций, связанных с клеточным дыханием. Это определяет характер и степень тяжести клинических проявлений митохондриалъных болезней.
Выдвинута также гипотеза о том, что накопление спонтанно возникающих мутаций мтДНК является звеном механизмов старения и развития дегенеративных процессов у человека.
Синдром MELAS (митохондриальная энцефаломиопатия, лактатацидоз, инсультоподобные эпизозы) Когнитивные расстройства различной степени и слабоумие, лактатацидоз, инсульты и переходные ишемические атаки
Пероксисомные болезни — группа заболеваний, обусловленных нарушением структуры и функции пероксисом, - внутриклеточных органелл, присутствующих в каждой клетке организма за исключением зрелых эритроцитов. Около 50 ферментов пероксисом обеспечивают Р-окисление очень длинноцепочечных жирных кислот (ОДЦЖК), дикарбоновых кислот, пипеколиновой кислоты, простагландинов. В пероксисомах происходят начальные этапы биосинтеза плазмалогенов, входящих в структуру миелина и составляющих от 5% до 20% фосфолипидов клеточных мембран. Одна из важнейших функций пероксисом — зашита клетки от образующегося в ней атомарного кислорода с помощью химических превращений, в которых участвуют пероксисомные каталазы.
Возникновение пероксисомных болезней обусловлено нарушением сложных ферментативных реакций внутри органелл, процессов транспорта белков через мембраны органелл и функции их рецепторов. Белки, вовлеченные в биогенез пероксисом, называют пероксинами. В настоящее время определены функции более 20 пероксинов, для 9 из которых установлена хромосомная локализация кодирующих генов, обозначаемых РЕХ. В большинстве случаев патогенетические механизмы пероксисомных болезней связаны с нарушением четырех основных биохимических процессов — синтеза плазмалогенов, окисления ОДЦЖК и фитановой кислоты и деградации пипеколиновой кислоты. Особенностью пероксисомных болезней является возникновение нескольких заболеваний при мутациях в одном и том же гене, или появление одного клинического фенотипа, при наличии мутаций в разных генах пероксинов. Среди наиболее распространенных пероксисомных болезней гепато-церебро-ренальный синдром Цельвегера, ризомелическаяточечная хондродиетрофия, синдром Рефсума и адренолейкодистрофия. Частота пероксисомных болезней составляет ): 25000-1:50000 новорожденных. На основании двух важных критериев — количества пероксисом в печени и степени нарушения их функций - эти болезни принято делить на три основные группы. К первой группе относятся пероксисомные болезни, при которых количество пероксисом в печени значительно снижено, а биохимические процессы в них нарушены. Во вторую группу входят заболевания, характеризующиеся нарушением лишь нескольких биохимических процессов в пероксисомах и нормальным их количеством в печени. При заболеваниях третьей группы биологическая функция пероксисом полностью подавлена, но их количество в печени соответствует норме. Клиническая картина пероксисомных заболеваний включает: 1) черспно-лицевые аномалии; 2) патологию органов зрения (нарушение пигментации сетчатки, побледнение дисков зрительных нервов); 3) неврологические симптомы (мышечная гипотония, судороги, задержка раннего психомоторного развития); 4) гепатомегалию; 5) аномалии скелета (ризомелический тип укорочения конечностей).
Для демонстрации особенностей проявления пероксисомных болезней приводим клинико-генетические характеристики церебро-гепато-ренального синдрома Цельвегера, относящегося к первой группе.
Синдром Цельвегера.
Синдром Цельвегера объединяет группу генетически гетерогенных состояний. К клиническим проявлениям синдрома Цельвегера могут приводить мутации в генах пероксинов 1,2,3,5,6 и 12. Все варианты СЦ наследуются по аутосомно-рецессивному типу.
Первые симптомы отмечаются с рождения. Для больных характерна внутриутробная гипотрофия (вес при рождении не превышает 2500 г), дисморфизм в строении лица и черепа — увеличение размеров лба, монголоидный разрез глаз, периорбитальная полнота тканей, короткий вздернутый нос, микрогнатия. Среди наиболее типичных признаков: резкая мышечная гипотония, доходящая до атонии, и поликистоз почек. У всех больных отмечаются полиморфные пороки развития головного мозга. Часто диагностируется полимикрогирия, лизэнцефалия, агенезия мозолистого тела, очаги демиелинизации в белом веществе мозга, гидроцефалия. В ряде случаев выявляется патология глаз в виде врожденных катаракт и глауком, а также пороки сердца и наружных половых органов. Для заболевания характерна длительная желтуха и симптомы надпочечниковой недостаточности в первые месяцы жизни. У всех детей отмечается грубая задержка раннего психомоторного развития и снижение продолжительности жизни. Большинство больных погибает в течение первого года.
Лизосомные болезни накопления – это тяжелые наследственные заболевания обмена веществ, связанные с отсутствием лизосомальных ферментов. Недостаток этих ферментов приводит к тому, что макромолекулы (сложные комплексы белков, липидов и углеводов) не расщипляются и накапливаются в лизосомах. В результате сначала нарушается работа, клетки, затем тканей, а затем всего организма. Частота заболеваний этой группой генетических болезней составляет 1:5000 новорожденных. В зависимости от самого субстрата и группы поврежденных ферментов различают: сфинголипидозы (ганглиозидоз, болезнь Крабе, болезнь Гоше, метахроматическая лейкодистрофия, , болезнь Фарбера, болезнь Фабри, , болезнь Шиндлера, болезнь Нимана-Пика); муколипидозы и гликопротеинозы (цероидный липофусциноз, болезнь Вольмана, муколипидоз маннозидоз); мукополисахаридозы (синдром Гурлера, синдром Хантера, синдром Шейе, синдром Сан-Филиппо, синдром Морото -_Лами, синдром Моркио, синдром Слая).
Диагноз лизосомного заболевания можно заподозрить на основе внешних признаков: скелетные аномалии, грубые черты лица, а также умственной отсталости, поражений внутренних органов и систем. Манифестация этих симптомов может произойти как в период новорожденности, так и в уже зрелом возрасте.
Одно из самых известных лизосомных заболеваний – болезнь Гоше. В основе лежит незаменимого фермента бета-глюкоцереброзидазы, в результате чего мембранный жир накапливается в клетках Гоша с нарушением функций внутренних органов.
Если раньше диагноз «Болезнь Гоше» считался практически приговором, то сейчас при применении заместительной ферментотерапии имиглюцеразой у больных появилась возможность достигнуть нормальной жизни. При регулярном приеме препарата размеры печени и селезенки уменьшаются практически до нормального состояния, гемограмма нормализуется, изчезают боли в костях.
ДНК – самые крупные биополимеры, содержащие до 108–109 мономеров – дезоксирибонуклеотидов, которые содержат сахар – дезоксирибозу. В состав ДНК входит 4 типа дезоксирибонуклеотидов: аденин – А, тимидин – Т, гуанин – G, цитозин – С.
АТФ (аденозинтрифософорная кислота, аденозинтрифосфат) – нуклеотид (рис. 28), образованный аденозином и 3 остатками фосфорной кислоты. Выполняет в организмах роль универсального аккумулятора энергии. Под действием ферментов фосфатные группы отщепляются от АТФ с освобождением энергии, благодаря которой происходят мышечные сокращения, синтетические и другие процессы жизнедеятельности.
В молекуле ДНК, состоящей из двух полинуклеотидных цепочек, выделяют первичную, вторичную, третичную и т.д. структуры.
Первичная структура представляет собой линейную последовательность дезоксирибонуклеотидов в одной цепочке. В такой форме в природе ДНК не существует, но именно первичная структура (последовательность нуклеотидов) определяет все ее свойства.
Стабильность. Она обеспечивается водородными, гликозидными и фосфодиэфирными связями, а также механизмом репарации спонтанных и индуцированных повреждений;
2. Способность к репликации. Благодаря этому механизму в соматических клетках сохраняется диплоидное число хромосом. Схематично псе перечисленные особенности ДНК как генетической молекулы изображены на рисунке. 3. Наличие генетического кода. Последовательность оснований в ДНК с помощью процессов транскрипции и трансляции преобразуется в последовательность аминокислот в полипептидной цепи;
4.Способность к генетической рекомбинации. Благодаря этому механизму образуются новые сочетания сцепленных генов.
Передача генетической информации в клетке основана на матричных процессах (репликации, транскрипции, трансляции). Синтез дочерней цепи (репликация) молекулы ДНК происходит по матрице одной из двух родительских цепей с образованием новой двухиепочечной молекулы ДНК. Синтез молекулы РНК совершается в процессе транскрипции ДНК по матрице одной из двух цепей ДНК. Такая матричная (информационная) РНК может рассматриваться как посредник между ДНК и белком. Далее при синтезе белков генетическая информация, закодированная в последовательности триплетов азотистых оснований (канонов), транслируется в аминокислотную последовательность полипептидных цепей. Остановимся кратко на каждом из этих процессов,
Репликация. Во время репликации происходит расхождение двух цепей ДНК, и каждая из них служит матрицей для синтеза дочерней цепи. Такой способ репликации называется полуконсервативным. При этом дезоксирибонуклеотиды встраиваются в дочернюю цепь согласно правилу комплементарности азотистых оснований (А — Т, G — С). Вновь образованная молекула состоит из одной родительской и одной дочерней цепи ДНК. Образование дочерних хромосом происходит на стадии синтеза (S) в интерфазе между митотическими делениями и перед первым делением мейоза, В анафазе удвоенные хромосомы расходятся по дочерним клеткам. Таким образом, без процесса репликации невозможно сохранение диплоидного числа хромосом в соматических клетках и образование гаплоидного набора хромосом в половых клетках после двух делений мейоза. Однако при делении клеток происходит не только сохранение числа хромосом, но и воспроизведение последовательности азотистых оснований в молекулах ДНК, основанное на комплементарностb пар оснований родительской и дочерней цепей ДНК.
Цепи отделяются друг от друга, и каждая служит матрицей для построения комплементарной цепи. В результате синтезируются две молекулы, у каждой из которых одна цепь старая и одна новая. Такой способ репликации ДНК называют полуконсервативным.
15.2.Поскольку ДНК является молекулой наследственности, то для реализации этого качества она должна точно копировать саму себя и таким образом сохранять всю имеющуюся в исходной молекуле ДНК информацию в виде определенной последовательности нуклеотидов. Это обеспечивается за счет особого процесса, предшествующего делению любой клетки ррганизма, который называется репликацией ДНК
Суть репликации днк заключается в том, что специальный фермент разрывает слабые водородные связи, которые соединяют между собой нуклеотиды двух цепей. В результате цепи ДНК разъединяются, и из каждой цепи «торчат» свободные азотистые основания (возникновение так называемой вилки репликации). Особый фермент ДНК-полимераза начинает двигаться вдоль свободной цепи ДНК от 5'- к З'-концу (лидирующая цепь), помогая присоединиться свободным нуклеотидам, постоянно синтезируемым в клетке, к З'-концу вновь синтезируемой цепи ДНК. На второй нити ДНК (отстающая нить) новая ДНК образуется в виде небольших сегментов, состоящих из 1000—2000 нуклеотидов
Для начала репликации днк фрагментов этой нити требуется синтез коротких фрагментов РНК (о характерных особенностях РНК будет сказано ниже) как затравок, для чего используется особый фермент — РНК-полимераза (праймаза). Впоследствии праймеры РНК удаляются, в образовавшиеся бреши встраивается ДНК с помощью ДНК полимеразы I. Таким образом, каждая цепь ДНК используется как матрица или шаблон для построения комплементарной цепи и репликация ДНК является полуконсервативной (т.е. одна нить в новой молекуле ДНК — «старая», а вторая — новая). Для репликации лидирующей и отстающей цепей клеткой используют разные ферменты. В результате репликации образуются две новые абсолютно идентичные молекулы ДНК, идентичные также исходной молекуле ДНК до начала ее редупликации (более подробно процесс репликации ДНК показан на рис. 3.5). ДНК-полимераза, как и любой другой фермент, существенно ускоряет процесс присоединения комплементарных нуклеотидов к свободной цепи ДНК, однако химическое сродство аденина к тимину, а цитозина к гуанину столь велико, что они соединяются друг с другом и в отсутствие ДНК-полимеразы в простой реакционной смеси1. Можно сказать, несколько упрощая, что феномен точного удвоения молекулы ДНК, в основе которого лежит компле-ментарность оснований этой молекулы, составляет молекулярную основу наследственности. Скорость репликации ДНК у человека относительно низкая и для того, чтобы обеспечить репликацию ДНК любой хромосомы человека, требовались бы недели, если бы репликация начиналась из одной точки. На самом деле в молекуле ДНК любой хромосомы, а-каждая хромосома человека содержит только одну молекулу ДНК, имеется множество мест инициации репликации (репликонов). От каждого репликона репликация идет в обоих направлениях до тех пор, пока соседние репликоны не сливаются. Поэтому репликация ДНК в каждой хромосоме протекает относительно быстро.
Полуконсервативная репликация. Две цепи исходной молекулы ДНК расходятся вследствие разрыва слабых водородных связей между азотистыми основаниями. Каждая из них служит матрицей для образования новой цепи ДНК, а возникающие между азотистыми основаниями водородные связи соединяют старую и новую цепи, восстанавливая целостность молекулы. В результате каждая новая клетка получает гибридную молекулу ДНК, состоящую из одной старой и одной новой цепи.
16.Классификация нуклеотидных последовательностей (уникальные иповторяющиеся ).В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1. Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2. Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3. Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие (
10 пн) некодирующие последовательности, которые входят в состав прицентромерного гетерохроматина.
роматин (греч.chroma — цвет, краска и греч.nitos — нить) — это вещество хромосом— комплексДНК,РНКибелков. Хроматин находится внутриядраклетокэукариоти входит в составнуклеоидаупрокариот. Именно в составе хроматина происходитреализация генетической информации, а такжерепликацияирепарация ДНК. Основную массу хроматина составляют белкигистоны. Гистоны являются компонентомнуклеосом, — надмолекулярных структур, участвующих в упаковке хромосом.
1. эухроматин, локализующийся ближе к центру ядра, более светлый, более деспирилизованный, менее компакт-ный, более активен в функциональном отношении. Эухроматин - неконденсированный хроматин, с которого происходит синтез белка.
2. Гетерохроматин - конденсированный хроматин, с которого белок не синтезируется. гетерохроматин - плотно спирализованная часть хроматина. Гетерохроматин соответствует конденсированным, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипции). Он интенсивно окра-шивается основными красителями, и в световом микроскопе имеет вид тёмных пятен, гранул.
Жизненный и митотический цикл клетки .Фазы митотического цикла ,их характеристика и значение.
Период существования клетки от момента ее образования путем деления материнской клетки (включая само деление) до собственного деления или смерти называют жизненным (клеточным) циклом.
Продолжительность жизненного цикла различных клеток многоклеточного организма различна. Так, клетки нервной ткани после завершения эмбрионального периода перестают делиться и функционируют на протяжении всей жизни организма, а затем погибают. Клетки же зародыша на стадии дробления, завершив одно деление, сразу приступают к следующему, минуя все остальные фазы.
Митоз – непрямое деление соматических клеток, в результате которого сначала происходит удвоение, а затем равномерное распределение наследственного материала между дочерними клетками.
Биологическое значение митоза: в результате митоза образуются две клетки, каждая из которых содержит столько же хромосом, сколько их было в материнской. Дочерние клетки генетически идентичны родительской. Число клеток в организме увеличивается, что представляет собой один из главных механизмов роста. Многие виды растений и животных размножаются бесполым путем при помощи одного лишь митотического деления клеток, таким образом, митоз лежит в основе размножения. Митоз обеспечивает регенерацию утраченных частей и замещение клеток, происходящее в той или иной степени у всех многоклеточных организмов.
Митотический цикл состоит из интерфазы и митоза. Длительность митотического цикла у разных организмов сильно варьирует. Непосредственно на деление клетки уходит обычно 1–3 ч, то есть основную часть жизни клетка находится в интерфазе.
Интерфазой называют промежуток между двумя клеточными делениями. Продолжительность интерфазы, как правило, составляет до 90% всего клеточного цикла. Интерфаза состоит из трех периодов: пресинтетический, или G1; синтетический, или S; постсинтетический, или G2.
Начальный отрезок интерфазы – пресинтетический период (2n2с, где n – количество хромосом, с – количество ДНК), период роста, начинающийся непосредственно после митоза. Синтетический период по продолжительности очень различен: от нескольких минут у бактерий до 6–12 ч в клетках млекопитающих. Во время синтетического периода происходит самое главное событие интерфазы – удвоение молекул ДНК. Каждая хромосома становится двухроматидной, а число хромосом не изменяется (2n4с).
Постсинтетический период. Обеспечивает подготовку клетки к делению и также характеризуется интенсивными процессами синтеза белков, входящих в состав хромосом; синтезируются ферменты и энергетические вещества, необходимые для обеспечения процесса деления клетки.
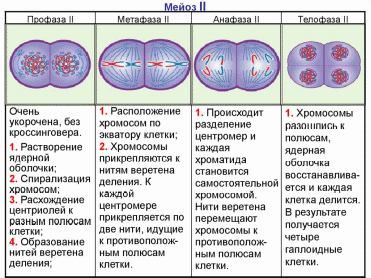
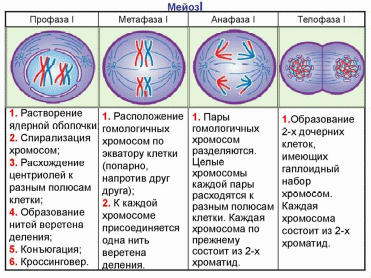
Митоз. Для удобства изучения происходящих во время деления событий митоз разделяют на четыре стадии: профазу, метафазу, анафазу, телофазу.
Профаза (2n4с). В результате спирализации хромосомы уплотняются, укорачиваются. В поздней профазе хорошо видно, что каждая хромосома состоит из двух хроматид, соединенных центромерой. Хромосомы начинают передвигаться к клеточному экватору. Формируется веретено деления, ядерная оболочка исчезает, и хромосомы свободно располагаются в цитоплазме. Ядрышко обычно исчезает чуть раньше.
Метафаза (2n4с). Хромосомы выстраиваются в плоскости экватора, образуя так называемую метафазную пластинку. Центромеры хромосом лежат строго в плоскости экватора. Нити веретена прикрепляются к центромерам хромосом, некоторые нити проходят от полюса к полюсу клетки, не прикрепляясь к хромосомам.
Анафаза (4n4с). Начинается с деления центромер всех хромосом, в результате чего хроматиды превращаются в две совершенно обособленные, самостоятельные дочерние хромосомы. Затем дочерние хромосомы начинают расходиться к полюсам клетки.
Телофаза (2n2с). Хромосомы концентрируются на полюсах клетки и деспирализуются. Веретено деления разрушается. Вокруг хромосом формируется оболочка ядер дочерних клеток, затем происходит деление цитоплазмы клетки (или цитокинез).
Репарация ДНК.
Процесс, позволяющий живым организмам восстанавливать повреждения, возникающие в ДНК, называют репарацией. Все репарационные механизмы основаны на том, что ДНК - двухцепочечная молекула, т.е. в клетке есть 2 копии генетической информации. Если нуклеотидная последовательность одной из двух цепей оказывается повреждённой (изменённой), информацию можно восстановить, так как вторая (комплементарная) цепь сохранена.роцесс репарации происходит в несколько этапов. На первом этапе выявляется нарушение комплементарности цепей ДНК. В ходе второго этапа некомплементарный нуклеотид или только основание устраняется, на третьем и четвёртом этапах идёт восстановление целостности цепи по принципу комплементарности. Однако в зависимости от типа повреждения количество этапов и ферментов, участвующих в его устранении, может быть разным.
Очень редко происходят повреждения, затрагивающие обе цепи ДНК, т.е. нарушения структуры нуклеотидов комплементарной пары. Такие повреждения в половых клетках не репарируются, так как для осуществления сложной репарации с участием гомологичной рекомбинации требуется наличие диплоидного набора хромосом.
А. Спонтанные повреждения
Нарушения комплементарности цепей ДНК могут происходить спонтанно, т.е. без участия каких-либо повреждающих факторов, напримерв результате ошибок репликации, дезаминирования нуклеотидов, депуринизации.
Ошибки репликации
Точность репликации ДНК очень велика, но примерно один раз на 105-106 нуклеотидных остатков происходят ошибки спаривания, и тогда вместо пары нуклеотидов А-Т, G-С в дочернюю цепь ДНК оказываются включёнными нук-леотиды, некомплементарные нуклеотидам матричной цепи. Однако ДНК-полимеразы δ, ε способны после присоединения очередного нук-леотида в растущую цепь ДНК делать шаг назад (в направлении от 3'- к 5'- концу) и вырезать последний нуклеотид, если он некомплементарен нуклеотиду в матричной цепи ДНК. Этот процесс исправления ошибок спаривания (или коррекция) иногда не срабатывает, и тогда в ДНК по окончании репликации остаются некомплементарные пары, тем более, что ДНК-полимераза а лишена корректирующего механизма и "ошибается" чаще, чем другие полимеразы.
При неправильном спаривании в первичной структуре дочерней цепи ДНК необычные основания не появляются, нарушена только ком-плементарность. Система репарации некомплементарных пар должна происходить только на дочерней цепи и производить замену некомплементарных оснований только в ней. Ферменты, участвующие в удалении неправильной пары нуклеотидов, распознают матричную цепь по наличию метилированных остатков аденина в последовательностях -GATC-.Пока основания нуклеотидных остатков в дочерней цепи неметилированы, ферменты должны успеть выявить ошибку репликации и устранить её.
Распознавание и удаление (первый этап) некомплементарного нуклеотида происходят при участии специальных белков mut S, mut L, mut H. Каждый из белков выполняет свою специфическую функцию. Mut S находит неправильную пару и связывается с этим фрагментом. Mut Н присоединяется к метилированному (по аденину) участку -GATC-, расположенному вблизи некомплементарной пары. Связующим между mut S и mut Н служит белок mut L, его присоединение завершает образование активного фермента. Формирование комплекса mut S, mut L, mut Н на участке, содержащем ошибку, способствует проявлению у белка mut Н эндонуклеазной активности. Ферментативный комплекс гидролизует фосфоэфирную связь в неметилированной цепи (рис. 4-21).
К свободным концам цепи присоединяется экзонуклеаза (второй этап). Отщепляя по одному нуклеотиду в направлении от 3'- к 5'- концу дочерней цепи, она устраняет участок, содержащий некомплементарную пару. Брешь застраивает ДНК-полимераза β (третий этап), соединение основного и вновь синтезированного участков цепи катализирует фермент ДНК-лигаза (четвёртый этап). Для успешного функционирования экзонуклеазы, ДНК-полимеразы р и ДНК-лигазы необходимо участие в репарации хеликазы и SSB-белков.
Б. Индуцируемые повреждения
Индуцируемые повреждения возникают в ДНК в результате воздействия разнообразных мутагенных факторов как радиационной, так и химической природы. Репарация необходима для сохранения нативной структуры генетического материала на протяжении всей жизни организма. Снижение активности ферментов репарационных систем приводит к накоплению повреждений (мутаций) в ДНК.
Причиной многих наследственных болезней человека выступает нарушение отдельных этапов процесса репарации.
Проблемы клеточной пролиферации в медицине.Определение пролиферативной активности клеток,тканей,органов.Значение метода тимидиновой радоавтографии в изучении жизненного цикла клетки.
Пролиферация (от лат.proles — отпрыск, потомство и fero — несу) — разрастание тканиорганизмапутёмразмножения клеток. Механизм пролиферации отличается от других механизмов изменения объёмаклетки(клеток), например,отёкаилиапоптоза. Термин в медицине впервые ввел немецкий ученыйВирховдля обозначения новообразования клеток путем их размножения делением. Регулировать интенсивность пролиферации можностимуляторамииингибиторами, которые могут вырабатываться и вдали от реагирующих клеток (например,гормонами), и внутри них. Непрерывно пролиферация происходит в раннемэмбриогенезеи по мере дифференцировки периоды между делениями удлиняются. Некоторые клетки, напримернервные, не способны к пролиферации.
С начала 60-х гг. появились новые взгляды на значение для старения и продолжительности жизни закономерностей клеточной пролиферации. На основании подсчета числа делений фибробластов, высеваемых в культуру ткани от эмбриона человека и от людей в возрасте 20 лет и выше, было сделано заключение о пределе клеточных делений(лимит Хейфлика), которому соответствует видовая длительность жизни. Показано, что фибробласты мыши способны удваивать свою численность 14—28 раз, цыпленка —15—35, человека—40—60, черепахи—72—114 раз. Проверка результатов, о которых идет речь, выявила, что представление об ограниченности числа клеточных делений в индивидуальном развитии является неточным.
Размножение.Формы и способы размножения.Половое размножение ,его эволюционное значение.
Размножение - это способность всех организмов воспроизводить себе подобных, что обеспечивает непрерывность и приемлемость жизни.
В основе бесполого размножения лежит деление клеток путем митоза, при котором из каждой материнской клетки (организма) создаются две равноценные дочерние клетки (два организма). Биологическая роль бесполого размножения заключается в возникновении организмов, идентичных родительским по содержанию наследственного материала, а также анатомическим и физиологическим свойствам (биологические копии).
Различают следующие способы бесполого размножения: деление, почкование, фрагментация, полиэмбриония, спорообразование, вегетативное размножение.эДеление — способ бесполого размножения, характерный для одноклеточных организмов, при котором материнская особь делится на две или большее количество дочерних клеток. Можно выделить: а) простое бинарное деление (прокариоты), б) митотическое бинарное деление (простейшие, одноклеточные водоросли), в) множественное деление, или шизогонию (малярийный плазмодий, трипаносомы). Во время деления парамеции (1) микронуклеус делится митозом, макронуклеус — амитозом. Во время шизогонии (2) сперва многократно митозом делится ядро, затем каждое из дочерних ядер окружается цитоплазмой, и формируются несколько самостоятельных организмов.
Почкование — способ бесполого размножения, при котором новые особи образуются в виде выростов на теле родительской особи (3). Дочерние особи могут отделяться от материнской и переходить к самостоятельному образу жизни (гидра, дрожжи), могут остаться прикрепленными к ней, образуя в этом случае колонии (коралловые полипы).
Фрагментация (4) — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается материнская особь (кольчатые черви, морские звезды, спирогира, элодея). В основе фрагментации лежит способность организмов к регенерации.
Полиэмбриония — способ бесполого размножения, при котором новые особи образуются из фрагментов (частей), на которые распадается эмбрион (монозиготные близнецы).
Вегетативное размножение — способ бесполого размножения, при котором новые особи образуются или из частей вегетативного тела материнской особи, или из особых структур (корневище, клубень и др.), специально предназначенных для этой формы размножения. Вегетативное размножение характерно для многих групп растений, используется в садоводстве, огородничестве, селекции растений (искусственное вегетативное размножение).
Спорообразование (6) — размножение посредством спор. Споры — специализированные клетки, у большинства видов образуются в особых органах — спорангиях. У высших растений образованию спор предшествует мейоз.
Клонирование — комплекс методов, используемых человеком для получения генетически идентичных копий клеток или особей. Клон — совокупность клеток или особей, произошедших от общего предка путем бесполого размножения. В основе получения клона лежит митоз (у бактерий — простое деление).
При половом размножении у прокариот две клетки обмениваются наследственной информацией в результате перехода молекулы ДНК из одной клетки в другую по цитоплазматическому мостику.
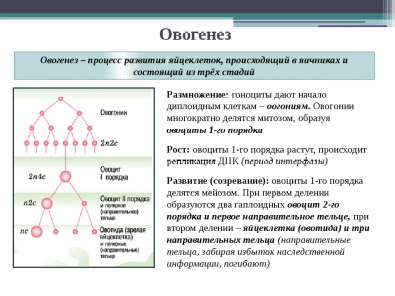
Половое размножение осуществляется при участии двух родительских особей (мужской и женской), у которых в особых органах образуются специализированные клетки — гаметы. Процесс формирования гамет называется гаметогенезом, основным этапом гаметогенеза является мейоз. Дочернее поколение развивается из зиготы — клетки, образовавшейся в результате слияния мужской и женской гамет. Процесс слияния мужской и женской гамет называется оплодотворением. Обязательным следствием полового размножения является перекомбинация генетического материала у дочернего поколения.
В зависимости от особенностей строения гамет, можно выделить следующие формы полового размножения: изогамию, гетерогамию и овогамию.
Изогамия (1) — форма полового размножения, при которой гаметы (условно женские и условно мужские) являются подвижными и имеют одинаковые морфологию и размеры.
Гетерогамия (2) — форма полового размножения, при которой женские и мужские гаметы являются подвижными, но женские — крупнее мужских и менее подвижны.
Овогамия (3) — форма полового размножения, при которой женские гаметы неподвижные и более крупные, чем мужские гаметы. В этом случае женские гаметы называются яйцеклетками, мужские гаметы, если имеют жгутики, — сперматозоидами, если не имеют, — спермиями.
Овогамия характерна для большинства видов животных и растений. Изогамия и гетерогамия встречаются у некоторых примитивных организмов (водоросли). Кроме вышеперечисленных, у некоторых водорослей и грибов имеются формы размножения, при которых половые клетки не образуются: хологамия и конъюгация. При хологамии происходит слияние друг с другом одноклеточных гаплоидных организмов, которые в данном случае выступают в роли гамет. Образовавшаяся диплоидная зигота затем делится мейозом с образованием четырех гаплоидных организмов. При конъюгации (4) происходит слияние содержимого отдельных гаплоидных клеток нитевидных талломов. По специально образующимся каналам содержимое одной клетки перетекает в другую, образуется диплоидная зигота, которая обычно после периода покоя также делится мейозом
У эукариот половой процесс связан с образованием половых клеток - ГАМЕТ. Мужскими гаметами являются сперматозоиды, женскими - яйцеклетки. Новый организм возникает в результате оплодотворения, СЛИЯНИЯ ЯДЕР ЯЙЦЕКЛЕТКИ И СПЕРМАТОЗОИДА. Образуется ЗИГОТА.
Очевидно, что гаметы должны иметь в два раза меньшее число хромосом, чем соматические клетки, так как в противном случае число хромосом в каждом последующем поколении должно было бы удваиваться. Этого не происходит благодаря особому типу клеточного деления МЕЙОЗУ.
При половом размножении в популяции создается более высокая генетическая изменчивость. В результате целого ряда процессов, гены, носителями которых изначально были родители, оказываются в новой комбинации в потомках. Именно благодаря рекомбинации внутри помета обнаруживаются многочисленные генетические различия, что повышает адаптационный потенциал популяции и вида в целом.
      |
|
|
 Скачать 1.14 Mb.
Скачать 1.14 Mb.