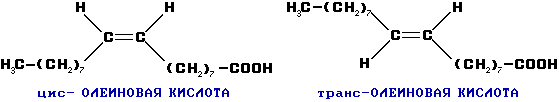Колоквиум по липидам. Дайте определение термину липиды
 Скачать 0.65 Mb. Скачать 0.65 Mb.
|

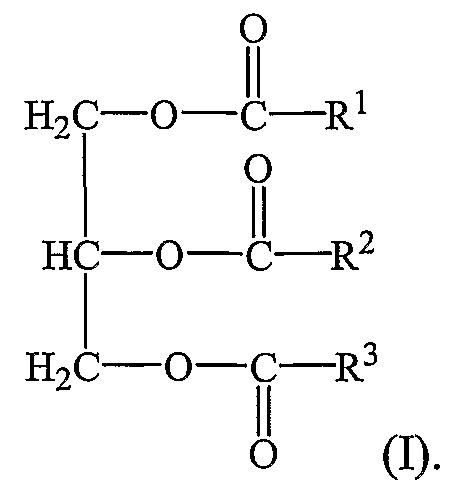
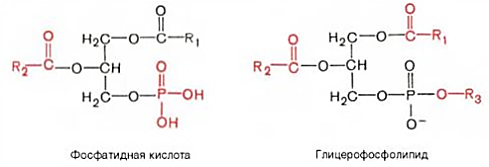
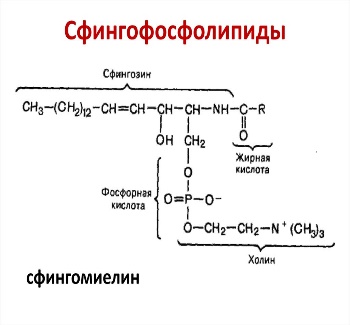
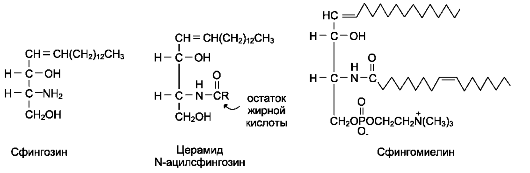
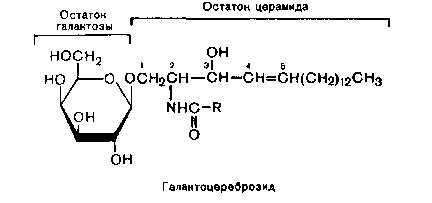
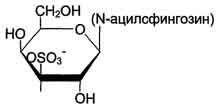
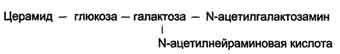
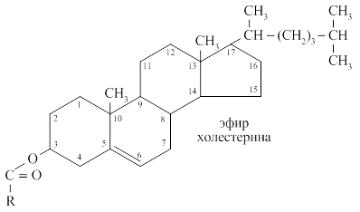
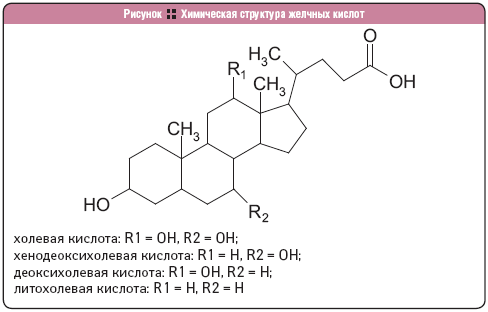
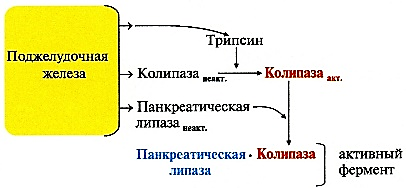
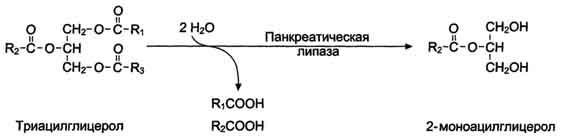
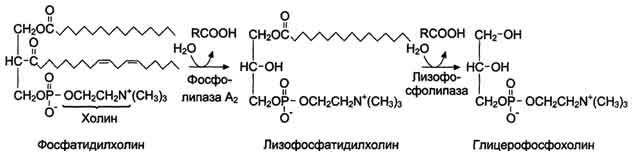
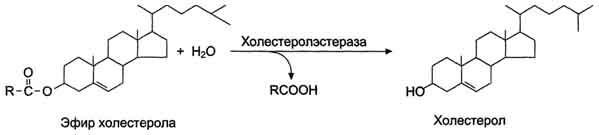
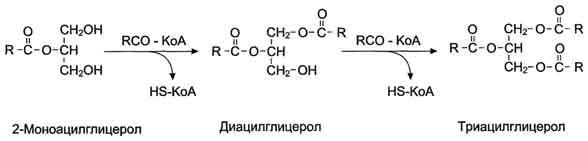
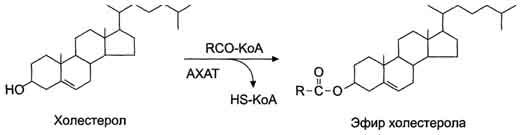
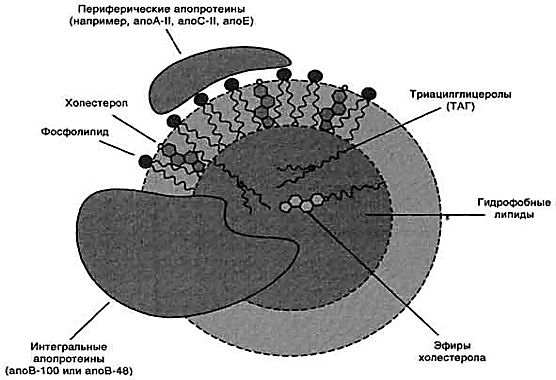
КоА + АМФ + PPi.Дайте определение термину «липиды». Изобразите на рисунке ненасыщенной жирной кислоты цис- и транс-изомерию. Назовите три известные вам насыщенные жирные кислоты. Назовите три известные вам ненасыщенные жирные кислоты. Что такое насыщенные жирные кислоты? Что такое ненасыщенные жирные кислоты? Объясните запись формулы жирной кислоты: C18:2Δ9,12. Объясните запись формулы жирной кислоты: C18:2Δω-6. Напишите общую структурную формулу триглицеридов. Напишите общую структурную формулу глицерофосфолипидов. Напишите общую структурную формулу сфингофосфолипидов. Напишите общую структурную формулу фосфатидной кислоты. Напишите общую структурную формулу сульфатидных цереброзидов. Перечислите витамины, которые поступают вместе с пищевыми жирами. Назовите основную функцию «липазы языка». Перечислите субстраты для синтеза глицерол-3-фосфата. Опишите последовательность событий при синтезе триглицеридов из жирных кислот и глицерол-3-фосфата в жировой ткани. Опишите последовательность событий при синтезе и секреции липопротеидов очень низкой плотности (ЛПОНП) в печени. Напишите реакцию, катализируемую триацилглицероллипазой (ТАГ-липазой) в адипоцитах. 20.продукты β-окисления жирных кислот. FADH2, NADH, ацетил-КоА 21.коферменты, необходимые для β-окисления жирных кислот. кофермент A, кофермент ФАДH2, кофермент Q !22.последовательность одного цикла реакций β-окисления жирных кислот. Ацил-SKOA = Еноил – SKOA = 3-гидроксиацил- SKOA = 3-кетоацил – SKOA = ацетил- SKOA. 23.регуляторы скорости β-окисления жирных кислот. скорость регулируется потребностью клетки в энергии, т.е. соотношениями АТФ/АДФ и NADH/NAD+,в тканях зависит от доступности субстрата, т.е. от количества жирных кислот, поступающих в митохондрии, ацетил-КоА-карбоксилаза, малонил-КоА 25.Рассчитайте количество АТФ, образующегося при β-окислении арахиновой кислоты (С20Н40O2). 120+45-2=163 26.Рассчитайте количество АТФ, образующегося при β-окислении стеариновой кислоты (C18H36O2). 148-2=146 27.Рассчитайте количество АТФ, образующегося при β-окислении пальмитиновой кислоты (C16H32O2). 131-2=129 28.Рассчитайте количество АТФ, образующегося при β-окислении миристиновой кислоты (C14H28O2). 114-2=112 29.Рассчитайте количество АТФ, образующегося при β-окислении лауриновой кислоты (C12H24O2). 97-2=95 30. три-четыре причины нарушения переноса жирных кислот в митохондрии. -длительный гемодиализ, в ходе которого организм теряет карнитин; длительная ацидурия, при которой карнитин выводится как основание с органическими кислотами, лечение больных сахарным диабетом препаратами сульфонилмочевины, ингибирующими карнитинацилтрансферазу I; низкая активность ферментов, синтезирующих карнитин; наследственные дефекты карнитинацил-трансферазы I. 31. последствия генетического дефекта дегидрогеназы среднецепочечных жирных кислот. Невозможность использовать жирные кислоты как источники энергии приводит к увеличению скорости окисления глюкозы. В результате у детей развивается гипогликемия - причина внезапной детской смертности (10% от общего числа умерших новорождённых). Если такие дети выживают, то после голодания в течение 6-8 ч у них развиваются гипогликемические приступы (слабость, головокружение, рвота, потеря сознания). Введение глюкозы приводит к исчезновению симптомов. 33.Опишите последствия нарушения окисления фитановой кислоты. фитановая кислота, поступающая с пищей, не окисляется и накапливается в организме, в основном в нервной ткани. Это приводит к нарушению структуры нервной ткани и развитию многих неврологических симптомов. 34. три соединения, называемых кетоновыми телами. -оксимасляную и ацетоуксусную кислоты и ацетон 35. функцию кетоновых тел. регуляторные функции, синтез АТФ, избавление от избытка кетоновых тел 37.Напиште реакцию синтеза В-гидроксибутирата из ацетоацетата. Последние 2 реакции. 38.Напишите реакцию синтеза Ацетоацетил-КоА из Ацетил-КоА. Первая реакция 39.Напишите реакцию,катализируемую ГМГ-КоА-Синтазой. Вторая реакция 40.Опишите регуляцию активности ГМГ-КоА-синтазы. 1.Гмг-КоА-синтаза- индуцируемый фермент,его синтез увеличивается при повышении концентрации жирных кислот в крови. 2.Ингибируется высокими концентрациями свободного кофермента А. 3.Когда поступление жирных кислот в клетки печени увеличивается, КоА связывается с ними, концентрация свободного КоА снижается, и фермент становится активным. 4.Если поступление жирных кислот в клетки печени уменьшается,то увеличивается концентрация свободного КоА, ингибирующего фермент. 44. «кетоацидоз». Кетоацидоз- накопление кетоновых тел в организме, которое приводит к уменьшению щелочного резерва, в тяжелых случаях- к сдвигу рН. 45. Назовите субстраты для синтеза жирных кислот в клетке. Ацетил-КоА в цитоплазме, оксалоацетат в цитозоле 48. Напишите суммарное уравнение синтеза пальмитиновой кислоты из ацетил-КоА и малонил-КоА. CH3-CO-SKoA + 7 HOOC-CH2-CO-SKoA + 14 (NADPH + H+) → C15H31COOH + 7 СО2+ 6 Н2О + 8 HSKoA + 14 NADP+. 1. ЖК называют полиеновыми? Ненасыщенные жирные кислоты содержащие 18 и более атомов углерода, с двумя и < числом двойных связей. 2.пример полиеновых ЖК Линолевая кислота, Арахидоновая кислота, α-линоленовая кислота, Эйкозапентаеновая кислота 3. Какие вещества называют эйкозаноидами? - биологически активные вещества , синтезируемые большинством клеток из полиеновых жк, содержащих 20 углеродных атомов. 4. класса эйкозаноидов. 1. простагландины;2. простациклины;3. тромбоксаны;(4. лейкотриены;) !5.общий для всех простагландинов структур элемент наличие циклопентанового кольца, 2 боковых цепей, двойной связи между C13- и С14-атомами и гидроксил группы у С15-атома !6. общий для всех тромбоксанов структурный элемент. тетрагидропирановый цикл. 8. стимула, активирующих фосфолипазы в клетке. Гормоны,гистамин,цитокины,механич воздействие 10. пример ингибитора фосфолипазы A₂. Глюкокортикоиды, Стероидные препараты 11. пример ингибитора циклооксигеназы. Аспирин и другие противовоспалительные препараты, ацетилирования серина в положении 530 в активном центре 12.Опишите различие свойств циклооксигеназы 1 и циклооксигеназы 2. Циклооксигеназа 1 - конститутивный фермент, синтезирующийся с постоянной скоростью. Синтез циклооксигеназы 2 увеличивается при воспалении и индуцируется соответствующими медиаторами - цитокинами. 14.примера эйкозаноида и их основного биологического действия. 1. PG E2 Расслабляет гладкую мускулатуру, расширяет сосуды, инициирует родовую активность, подавляет миграцию лимфоцитов, пролиферацию Т-клеток.2. PG F2α Сокращает гладкую мускулатуру, суживает сосуды, бронхи, стимулирует сокращения матки.3. PG D3 Вызывает расширение сосудов, снижает агрегацию тромбоцитов и лейкоцитов. 15. схему процесса инактивации эйкозаноидов. Простагландины инактивируются путём окисления гидроксильной группы в положении 15, важнейшей для их активности, до кетогруппы. Двойная связь в положении 13 восстанавливается. Затем происходит β-окисление боковой цепи, а после него - ω-окисление. Конечные продукты (дикарбоновые кислоты) выделяются с мочой. Активный ТХ A2 быстро превращается в биологически неактивный IX В2 путём разрыва кислородного мостика между 9 и 11 атомами углерода с образ гидроксильных групп. 16. осн ф-ю сурфактанта в респиратор отделе легких Слой сурфактанта покрывает внутр п-ть стенки альвеол и состоит из 2 фаз — поверхностной мембранной (апофаза), представленной молекулярной пленкой фосфолипидов, и жидкой (гипофаза), содержащей липиды, белки, полисахариды, воду и др., которая заполняет неровности и пространства между эпителиоцитами. 18 функции гликосфинголипидов. 1.Взаимодействие между: клетками, клетками и межклеточным мартиксом , клетками и микробами.2.Модуляция: активности протеинкиназ, активности рецепрора фактора роста, антипролиферативного действия.3.Обеспечение: структурной жесткости мембран, конформации белков мембран 20. пример заболевания, обусловленного дефектом лизосомальных ферментов катаболизма гликосфинголипидов. болезнь Гоше вследствие дефекта фермента β-глюкрзидазы, болезнь Тея-Сакса (дефект фермента β-гексозаминидазы) 21. функции холестерина укрепляет стенки клетки; производит желчную к-ту, которая расщепляет жиры в кишечнике; спос-ет образ в организме вит D и гормонов: тестостерон у муж и эстроген у жен 24. механизм регуляции ГМГ-КоА-редуктазы инсулином. Действие инсулина осуществляется через 2 фермента: фосфатазу киназы ГМГ-КоА-редуктазы, которая превращает киназу в неактивное дефосфорилированное состояние; фосфатазу ГМГ-КоА-редуктазы путём превращения её в дефосфорилированное активное состояние. Результатом этих реакций служит образование дефосфорилированной активной формы ГМГ-КоА-редуктазы. 25. механизм регуляции ГМГ-КоА-редуктазы глюкагоном. глюкагон через протеинкиназу А стимулирует фосфорилирование ГМГ-КоА-редуктазы, переводя её в неактивное состояние. Действие усиливается ,т.к. одновременно глюкагон стимулирует фосфорилирование и инактивацию фосфатазы ГМГ-КоА-редуктазы и фосфорилирование киназы ГМГ-КоА-редуктазы, удерживая, таким образом, ГМГ-КоА-редуктазу в фосфорилированном неактивном состоянии. В результате синтез холестерола в постабсорбтивном периоде и при голодании ингибируется. 26. транспортные формы холестерина Транспорт экзогенного холестерола, Тр-т эндогенного холестерола в сост ЛПОНП (пре-β-липопротеинов), Транспорт холестерола в составе ЛПНП. 27. цикл работы рецептора ЛПНП в клетке. Рецепторы ЛПНП синтезируются в ЭР и аппарате Гольджи, а затем экспонируются на поверхности клетки, в специальных углублениях, выстланных белком клатрином. Эти углубления называют окаймлёнными ямками. Выступающий на поверхность N-концевой домен рецептора взаимодействует с белками апоВ-100 и апоЕ; поэтому он может связывать не только ЛПНП, но и ЛППП, ЛПОНП, остаточные ХМ, содержащие эти апопротеины. Клетки тканей содержат большое количество рецепторов ЛПНП на своей поверхности: например, на одной клетке фибробласта имеется от 20 000 до 50 000 рецепторов 28. механизм влияния холестерола на собственный синтез и захват клеткой из крови. ГМГ-КоА-редуктазы в печени трансформируют в активную форму и эффективно ингибируют регуляторный фермент биосинтеза холестерола. Такие препараты могут практически полностью подавить синтез собственного холестерола в организме. В этих условиях печень увеличивает захват холестерола из крови. Для этого в клетках печени почти вдвое увеличивается синтез белков-рецепторов Л ПНП и, соответственно, увеличивается захват ЛПНП из крови. 29. цикл работы ЛПВП при обратном транспорте холестерина в печень. Незрелые ЛПВП-предшественники обогащаются холестеролом, который поступает в ЛПВП при участии фермента ЛХАТ с поверхности клеток и других липопротеинов, содержащих холестерол. Незрелые ЛПВП, обогащаясь холестеролом, превращаются в ЛПВП3 - частицы сферической формы и большего размера. ЛПВП3 обменивают эфиры холестерола на триацилглицеролы, содержащиеся в ЛПОНП, ЛППП при участии "белка, переносящего эфиры холестерола". ЛПВП3 превращается в ЛПВП2, размер которых увеличивается за счёт накопления триацилглицеролов. ЛПОНП и ЛППП под действием ЛП-липазы превращаются в ЛПНП, которые доставляют холестерол в печень. Часть ЛПВП захватывается клетками печени, взаимодействуя со специфическими для ЛПВП рецепторами к апоА-I. На поверхности клеток печени фосфолипиды и триацилглицеролы ЛППП, ЛПВП2 гидролизуются печёночной ЛП-липазой, что дестабилизирует структуру поверхности ЛП и способствует диффузии холестерола в гепатоциты. ЛПВП2 в результате этого опять превращаются в ЛПВП3 и возвращаются в кровоток. 31. общую схему синтеза конъюгатов желчных кислот. Конъюгация происходит в клетках печени и начинается с образования активной формы жёлчных кислот - производных КоА. Затем присоединяется таурин или глицин, и в результате образуется 4 варианта конъюгатов: таурохолевая и таурохенодезоксихолевая, гликохолевая или гликохенодезоксихолевая кислоты .Конъюгатов с глицином образуется в 3 раза больше, чем с таурином, так как количество таурина ограничено. 33. механизм регуляции синтеза желчных кислот. Регуляторные ферменты синтеза жёлчных кислот (7-α-гидроксилаза) и холестерола (ГМГ-КоА-редуктаза) ингибируются жёлчными кислотами. В течение суток активность обоих ферментов меняется сходным образом, т.е. увеличение количества жёлчных кислот в печени приводит к снижению синтеза как жёлчных кислот, так и холестерола. Возвращение жёлчных кислот в печень в процессе энтерогепатической циркуляции оказывает важное регуляторное действие; прерывание циркуляции приводит к активации 7-α-гидроксилазы и увеличению захвата холестерола из крови. Этот механизм лежит в основе одного из способов снижения конценграции холестерола в крови при лечении гиперхолестеролемии. В этом случае используют препараты, адсорбирующие в кишечнике холестерол и жёлчные кислоты и препятствующие их всасыванию. 34. причины генетической дислипопротеинемии. Дислипопротеинемии проявляются чаще всего повышением концентрации либо одного типа ЛП, либо сочетанным увеличением содержания нескольких типов ЛП. 36. Опишите механизм действия статинов на синтез холестерина. |