Документ Microsoft Word. Длина 24см Ширина 12см
 Скачать 195.69 Kb. Скачать 195.69 Kb.
|

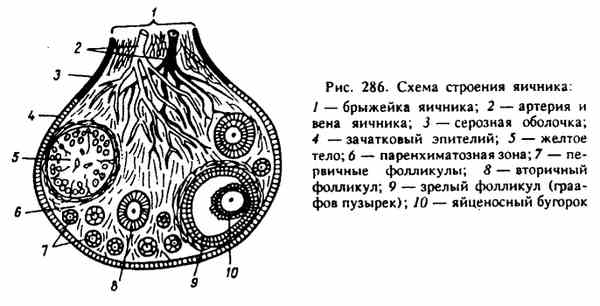 Среди высших позвоночных птицы имеют самый высокий репродуктивный потенциал. Так, масса яиц, снесенных курицей-несушкой за год, превышает массу ее тела в 10...22 раза. Высокая продуктивность, связанная с интенсивным обменом веществ, небольшие размеры и значительные адаптационные возможности сделали птицу одним из наиболее рентабельных объектов современного промышленного животноводства. Половое созревание курицы яйценоских пород завершается к 4,5 мес, более крупных мясных — к 6...8 мес; петушки созревают несколько раньше. Продолжительность половой активности домашней птицы составляет у петуха до 4 лет, кур до 6, у индюка до 3, у индейки до 5, у селезня и утки до 4, у гусака до 5, а у гусыни до 8 лет. Морфология гонад птиц принципиально не отличается от их строения у многих позвоночных. Семенники у самцов птиц расположены в брюшной полости на уровне трех последних ребер. Это парные органы, одетые соединительнотканной оболочкой, от которой вглубь семенника отходят прослойки, разделяющие его на дольки. Внутри долек находятся извитые семенные канальцы. Канальцы снаружи покрыты базальной мембраной, к которой примыкают крупные клетки Сертоли, которые выполняют трофические функции. Между клетками Сертоли расположены клетки сперматогенного эпителия на разных стадиях развития. Между канальцами находится рыхлая соединительная ткань, содержащая кровеносные сосуды и крупные клетки Лейдига. Особенности семенников у птиц — значительные размеры и большое число семенных канальцев, относительно малое количество рыхлой соединительной ткани, тенденция к редукции правого семенника и др. В период половой деятельности семенники становятся крупнее; в извитых канальцах образуются спермии. Спермии через семявыносящие канальцы, которые впадают в придаток семенника, переходят в семяпровод, открывающийся в стенке клоаки сосочком. Придатки семенников развиты слабо, придаточные половые железы у птиц отсутствуют. Совокупительного органа у петуха нет, он вводит семя, прижимая свою клоаку к клоаке самки. У селезней и гусаков копулятивный орган (половой член) состоит из фиброзной ткани, образующей кавернозное тело. На его поверхности спирально проходит семенной желоб, который при эрекции превращается в канал. Объем эякулята у петуха достигает 0,8 мл, количество спермиев в 1 мл составляет 5...7 млрд, при частом совокуплении количество спермиев снижается. С наступлением линьки половая активность петуха постепенно ослабевает и наступает состояние полового покоя. В отличие от яйцеклеток спермии способны длительное время сохраняться в яйцеводе кур, не теряя своей оплодотворяющей способности. Например, куры несут оплодотворенные яйца еще в течение 20 сут после отсадки самцов, у индеек и гусынь этот срок более продолжителен. У регулярно спаривающихся кур все отделы яйцевода заполнены спермиями. После спаривания спермии не сразу появляются в воронке яйцевода, где происходит оплодотворение. Первые оплодотворенные яйца курица несет лишь на 3 сут. У самок птиц функционирует только левый яичник; правый, хотя и закладывается, вскоре редуцируется. Соответственно у них имеется только один яйцевод, состоящий из воронки, белковой части, перешейка, матки («известковой» части) и влагалища, которое открывается в клоаку. Общая длина яйцевода у кур-несушек достигает 60 см. Яичник подвешен на брыжейке под передней долей почки. В яичнике различают корковое и мозговое вещество. Поверхность яичника покрыта однослойным эпителием. Под ним находится соединительнотканная оболочка, за которой расположены два слоя яйцевых фолликулов. В поверхностном слое лежат мелкие фолликулы, в глубоком — более крупные и зрелые. Мозговое вещество, расположенное в центре яичника, состоит из соединительной ткани, содержащей большое число кровеносных сосудов и нервных стволов. В нем имеются лакуны, выстланные плоским однослойным эпителием. Его глубокий слой образует сосудистую зону, поверхностный слой — фолликулы разной величины и зрелости, прикрепленные на богатых кровеносными сосудами выростах серозной оболочки. В разгар яйцекладки яичник напоминает кисть винограда; у курицы-несушки в нем содержится от 500 до 3500 ооцитов на разных стадиях оогенеза. Первое мейотическое деление происходит примерно за 1 ч до овуляции. Завершение второго мейотического деления и выделение второго направительного тельца происходят уже после разрыва фолликула, попадания яйца в воронку яйцевода и проникновения в яйцо спермия. В фолликуле различают ряд слоев клеток, окружающих ооцит. Ближе всего к ооциту лежат радиальная и гранулярная зоны, далее следует соединительнотканная оболочка фолликула, состоящая из базальной мембраны и собственно теки (внешней и внутренней). Зрелые фолликулы, висящие на ножке в полости тела, дополнительно покрыты соединительнотканной оболочкой яичника и эпителием. На апикальном полюсе фолликула оболочки утончаются. Это место разрывается при овуляции, его называют стигмой. Все соединительнотканные оболочки фолликула, находящиеся снаружи от базальной мембраны, содержат значительное число гладких мышечных волокон, хорошо васкуляризированы и иннер-вированы. Характерные особенности яичников птиц: отсутствие полости фолликула, огромные размеры ооцита (за счет запаса желтка), гипертрофия левого и редукция правого яичника. Гонады у птиц, как и у других позвоночных, выполняют две основные функции — продукцию гамет и секрецию половых гормонов. Гаметогенез подразделяется на ряд стадий: размножение, малый и большой (у самок) рост и созревание. Стадия размножения, во время которой формируется основной пул половых клеток, обычно заканчивается к моменту вылупления. Малый (цитоплазматический) рост завершается к периоду половой зрелости. В стадию большого роста (накопления желтка в ооците) фолликулы вступают за несколько дней до овуляции, причем вступление разных ооцитов в эту стадию происходит не одновременно, а, как правило, с интервалом в 1 сут (явление фолликулярной иерархии). Часть овариальных фолликулов птиц, как и у млекопитающих, подвергается атрезии. Деление созревания (мейоз) у самцов начинается перед половым созреванием и имеет место во время всего репродуктивного периода. У самок первое деление созревания происходит непосредственно перед овуляцией, а второе — уже после разрыва стигмы, овуляции и попадания яйцеклетки в яйцевод. В ходе своего развития ооциты накапливают в цитоплазме желток, за счет которого достигают значительных размеров (35...40 мм в диаметре у курицы). В созреваюших яйцах выделяют первичный и вторичный желток. Первичный желток является производным цитоплазматических структур самого ооцита, а вторичный — результатом секреторной деятельности клеток фолликулярного эпителия. Образование желтка длится около 2 нед. Желток куриного яйца состоит из мелких и крупных гранул; более крупные из них называются желточными шарами. Различают белый и желтый желток по интенсивности окраски и строению желточных шаров. Слои белого и желтого желтков чередуются. В процессе развития ооцита в яичнике курицы слои желтого желтка откладываются в цитоплазме ооцита днем, а слои белого желтка — ночью, когда в яичник поступает меньше питательных веществ из крови. 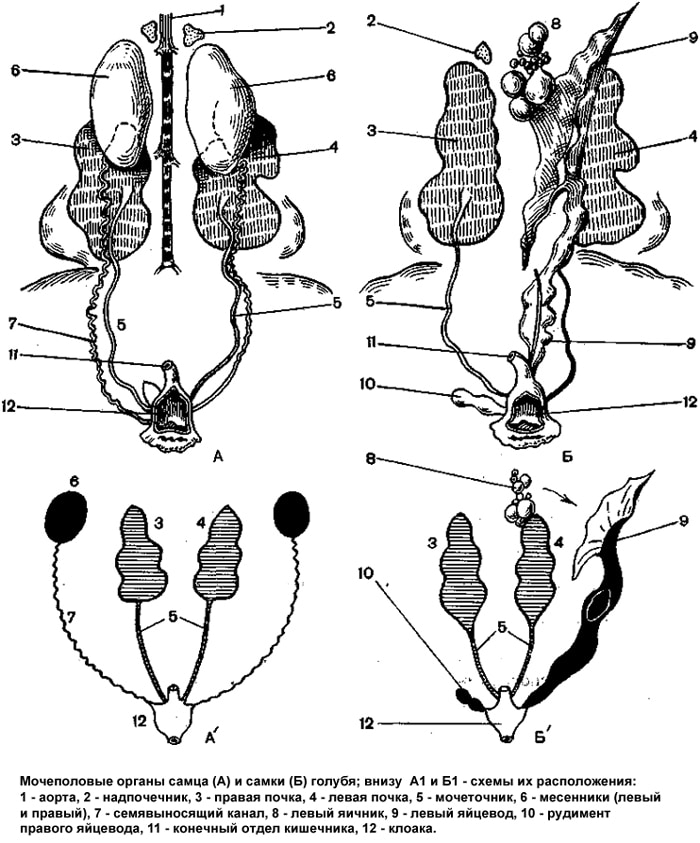 Половой диморфизм Наиболее отчетлив половой диморфизм у куриных, гусеобразных и воробьиных, у которых самцы заметно крупнее самок, более ярко окрашены, иногда имеют более громкий голос и сложную песню. У других видов птиц он выражен только в размерах: самцы несколько крупнее самок. У дневных хищных птиц и сов самки крупнее самцов. Половые различия в окраске характерны в отдельных случаях не только для оперения, но и для других частей тела – клювы, радужины, голых частей кожи. Иногда половые различия выражены в наличии у одного пола (обычно у самцов) выростов и придатков кожи на голове (у куриных), в присутствии шпор у самцов (у фазановых), в развитии характерных перьев – хохлов, длинных рулевых перьев (у фазанов и др. ), в устройстве голосового аппарата (в основном у утиных и певчих воробьинообразных), в пропорциях и размерах отдельных частей тела (клюв часто крупнее у самца, таз относительно шире у самок) и т. д. Половой диморфизм проявляется обычно у птиц, достигших половой зрелости. Он играет важную роль в нормальном течении полового цикла. Внешние различия между полами облегчают опознание и нахождение брачного партнера. Половой диморфизм играет важную роль в половом отборе. Согласно одной из теорий, половой диморфизм выражен тем больше, чем различнее вклады обоих полов в уход за потомством. Половой диморфизм отсутствует у некоторых систематических групп птиц (у пингвинов, трубконосых, гагар, стрижей и ряда других). Очень слабо выражен он у чаек, чистиков, пастушков, куликов и многих воробьиных. Анатомия половой системы самцов Морфология гонад птиц принципиально не отличается от их строения у многих позвоночных. Парные бобовидные семенники подвешены на брыжейке около переднего края почек. К началу размножения по сравнению с периодом покоя их объем возрастает в 300 -1000 раз. К внутреннему краю каждого семенника прилегает придаток семенника - остаток мезонефрической почки. От каждого придатка семенника начинается тонкий семяпровод (гомологичен вольфову каналу), проходящий по брюшной поверхности почки и впадающий в клоаку, образовав перед этим небольшое расширение - семенной пузырек - резервуар, где скапливаются зрелые сперматозоиды. Копулятивные органы в виде непарного выворачивающегося участка стенки клоаки есть лишь у немногих птиц - страусов, тинаму, гусеобразных. У остальных птиц оплодотворение происходит прижимании наружного отверстия клоаки самца к клоаке самки. Анатомия половой системы самок У самок развивается только левый яичник; редукция правого яичника (и правого яйцевода), вероятно, определяется тем, что при крупных размерах яиц одновременное их формирование в парных яичниках невозможно. Яичник подвешен на брыжейке около переднего конца левой почки. Левый яйцевод (мюллеров канал) воронкой открывается в полость тела около яичника, а его утолщенный задний (маточный) отдел впадает в левую часть клоаки. У части самок встречается небольшой слепой вырост клоаки - остаток редуцированного правого яйцевода. К началу размножения часть фолликулов увеличивается в размерах, так как находящиеся в них ооциты интенсивно накапливают желток. Одновременно происходит удлинение яйцевода и набухание его стенок. Созревшая яйцеклетка через разрыв стенки фолликула выпадает в полость тела и, подхваченная сильно увеличившейся воронкой, попадает в яйцевод. Оплодотворение должно произойти в начальной части яйцевода, примыкающей к воронке. После оплодотворения яйцеклетки происходит образование ее третичных оболочек: белка, двух подскорлуповых оболочек, скорлупы. Поэтому строение стенки яйцевода изменяется в разных его зонах. Яйцевод состоит из воронки, белкового отдела, перешейка, матки, влагалища. Стенка всех этих отделов построена из слизистой, мышечной и серозной оболочек. После этого яйцеклетка сокращениями стенок яйцевода перемещается по направлению к клоаке, покрываясь несколькими оболочками - выделениями желез стенок яйцевода. От момента проникновения яйцеклетки в яйцевод до полностью сформированного и готового к откладке яйца у разных птиц проходит 12 - 48 ч. В период яйцекладки число фолликулов достигает более 900, между яйцекладками их становится значительно меньше и они имеют микроскопические размеры. После овуляции фолликулярный эпителий остается прикрепленным к стенке опустевшего фолликула. Желтое тело не образуется. Строма мозгового вещества богата кровеносными сосудами. В нем находятся медулярные интерстициальеые клетки и лакуны, покрытые однослойным эпителием. |