Ответы на теоритеческие вопросы экзамен по физиологии. Друга група місцевих міогенних механізмів регуляції тонусу судин
 Скачать 104.63 Kb. Скачать 104.63 Kb.
|

|
Дихання 125. Загальна характеристика системи дихання. Основні етапи дихання. Біомеханіка вдіху и відіху. Дихання людини забезпечує дихальна система, що складається з повітроносних верхніх (носова порожнина, носоглотка, ротоглотка) і нижніх (гортань, трахея і бронхи) шляхів та легень Процеси дихання прийнято розділяти на 5 етапів 1) вентиляція легень транспорт газів із зовнішнього середовища в альвеоли і в зворотному напрямку та обмін газами між повітроносними шляхами та альвеолами у транзиторній зоні бронхіального дерева; 2) обмін дихальними газами між альвеолами і кров’ю 3) транспорт дихальних газів кров’ю 4) обмін дихальними газами між кров’ю і тканинами 5) внутрішнє (тканинне) дихання-процеси біологічного окислення органічних субстратів, які відбуваються в мітохондріях клітин Механізм вдиху. Вдих починається з активації інспіраторних нейронів дихального центру. Імпульси від них по tr. reticulospinalis передаються у спинний мозок на Механізм видиху. Імпульси не надходять до інспіраторних м'язів, вони розслаблюються, об'єм грудної порожнини зменшується. Об'єм плевральної порожнини зменшується, плевральний тиск зростає (-3-5 см Н2О). Транспульмональний тиск зменшується, об'єм альвеол зменшується, тиск в них зростає: він стає більшим за атмосферний. І за градієнтом тиску повітря із альвеол виходить в атмосферу. 126. еластичності тяга легень, негативний Тиск у плевральній щіліні. Еластична тяга легень – це сила, з якою легені прагнуть займати найменший об'єм. Вона зумовлена такими чинниками: 1) еластичним натягом легеневої тканини; 2) поверхневим натягом молекул рідини, що вкриває внутрішню поверхню альвеол (2/3 усієї сили); 3) тонусом бронхіальних м'язів. На поверхні легень утворюється напруга, зумовлена розтягом еластичних і колагенових волокон, вплетених в їх паренхіму, які весь час прагнуть скоротитись і зменшити об'єм органа (еластична тяга). Однак стисненню легень перешкоджає негативний плевральний тиск, спрямований на їх розправлення . У Межплевральное щілини знаходиться невелика кількість серозної рідини. При вдиху об'єм грудної порожнини зростає. А так як плевральна ізольована від атмосфери, то тиск в ній знижується. Легкі розширюються, тиск в альвеолах стає нижче атмосферного. Повітря через трахею і бронхи надходить в альвеоли. Під час видиху об'єм грудної клітини зменшується. Тиск в плевральній щілини зростає, повітря виходить з альвеол. Руху або екскурсії легких пояснюються коливаннями негативного Межплевральное тиску. Після спокійного видиху воно нижче атмосферного на 4-6 мм.рт.ст. На висоті спокійного вдиху на 8-9 мм.рт.ст. Після форсованого видиху воно нижче на 1-3 мм.рт.ст., а форсованого вдиху на 10-15 мм. рт. ст. 127. Зовнішнє дихання. Показники зовнішнього дихання та їх оцінка. ЗОВНІШНЄ, АБО ЛЕГЕНЕВЕ ДИХАННЯ – обмін газів між атмосферним повітрям і організмом. Легеневе дихання – це дихання за допомогою внутрішніх спеціалізованих органів газообміну (легенів) у тісному взаємозв'язку дихальної системи з кровоносною, опорно – руховою та регуляторними системами. До статичних належать такі об'єми. 1Дихальний об'єм (ДО) - це об'єм повітря, який доросла людина в стані спокою вдихає і видихає (приблизно 500 мл, коливання від 300 до 800 мл). 2 Резервний об'єм вдиху (РОвд) – це додатковий об'єм повітря, який людина може вдихнути при глибокому вдиху після спокійного видиху (3000 мл). 3 Резервний об'єм видиху (РОвид) – максимальний додатковий видих повітря після спокійного видиху (1300 мл). 4 Залишковий об'єм (30) - об'єм повітря, що залишається в легенях після максимального видиху (1200 мл), (рис. 11.8). 5 Мертвий простір (МП) - об'єм повітря, що знаходиться в повітроносних шляхах і не бере участі в газообміні (150 мл). До легеневих ємностей належать: Ємність вдиху (Є ) - це об'єм повітря, який може вдихнути людина після спокійного видиху. Він складається із ДО (500 мл) і РОвд (3000 мл) і становить в середньому 3500 мл. 2 Життєва ємність легень (ЖЄЛ) - максимальний об'єм повітря, який людина може видихнути після максимально глибокого вдиху. Це сума об'ємів, що складається із ДО (500 мл), РОвд (3000 мл) та РОвид (1300 мл) і становить 4800 мл. ЖЄЛ є об'єктивним показником, що свідчить про об'єм повітря, яке може бути введене і виведене із легень за один дихальний цикл. У чоловіків вона може сягати 5500 мл, у жінок - 4500 мл. Залежить від віку, фізичного розвитку, росту та маси тіла людини. 128. анатомічний и фізіологічній "мертвий простір", его фізіологічна роль. Повітря, яке знаходиться в повітряносних шляхах не бере участь в газообміні називається «анатомічний мертвий простір»,він складає 150 мл. Наприклад, людина вдихнула 500мл,але лише 350 мл. беруть участь у газообміні в альвеолах .Анатомічний мертвий простір відіграє позитивну роль, бо повітря в повітроносних шляхах зігрівається,очищується, зволожується. Альвеолярний простір – це сукупність альвеол, які в дану мить не беруть участь в газообміні, бо не наповнюються повітрям або навколо них неефективний капілярний кровообіг. Сума анатомічного і альвеолярного мертвого простору називається фізіологічним мертвим простором. 129. дифузія газів у легень. Діфузійна здатність легень и фактори, від якіх вона покладу. Цим терміном позначається кількість газу, яка проходить через легеневу мембрану за 1 хв при градієнті тиску 1 мм рт. ст. Для кисню цей показник складає 25–З0 мл/хв мм рт. ст. З віком, як і при ряді патологічних станів, цей показник знижується. Це відбувається через зменшення обмінної площі альвеол, збільшення відстані при дифузії у разі набряку чи склеротичних змін. Ефективність газообміну у легенях залежить від швидкості кровотоку. Еритроцит проходить по капіляру за 0,6-1 с. За цей час РАО2, і РаО2 встигають вирівнятися. Але при надмірному підвищенні швидкості кровотоку, наприклад при інтенсивному фізичному навантаженні, еритроцит через легеневий капіляр може "проскакувати" швидше, ніж за критичні 0,2-0,25 с, і тоді насичення крові киснем знижується. 130. Транспорт кисня кров'ю. Киснева Ємність крови. Кисень транспортується кров’ю у двох формах: 1. Розчинений у плазмі крові. При РСО2 =100 мм.рт.ст. в 1л крові розчиняється 3 мл кисню. 2. В хімічно зв’язаному з гемоглобіном стані – у вигляді оксигемоглобіну. Це основна форма транспорту кисню – 1г гемоглобіну за оптимальних умов може зв’язати 1,34 мл кисню. Виходячи з цього розраховують кисневу ємкість крові – максимальну кількість О2 , котру може зв’язати 1л крові. Киснева ємність крові – це максимальна кількість кисню, яка може бути зв'язана зі 100 мл крові. Вона становить 20 мл (у венозній крові 12-16 мл). Кількість O2 у крові зумовлена кількістю розчиненого O2, кількістю гемоглобіну в крові й спорідненістю гемоглобіну до O2. 131. Крива дісоціації оксігемоглобіну, фактори, что вплівають на ее Хід. Крива дисоціації оксигемоглобіну –нелінійне співвідношення у відсотках O2-транспортувальної сили насичення гемоглобіну киснем до напруги кисню; має S-подібну форму На криву дисоціації оксигемоглобіну впливають: pH, температура і концентрація 2,3-дифосфогліцерату (2,3-ДФГ). Підвищення температури, зростання концентрації 2,3-ДФГ, зменшення pH зміщує криву праворуч, тобто, за тієї ж напруги кисню оксигемоглобіну утворюється менше (рис 8.4).551'552 Зручним показником для таких зміщень є Р50 – напруга кисню (РO2), за якої гемоглобін насичується O2 наполовину. 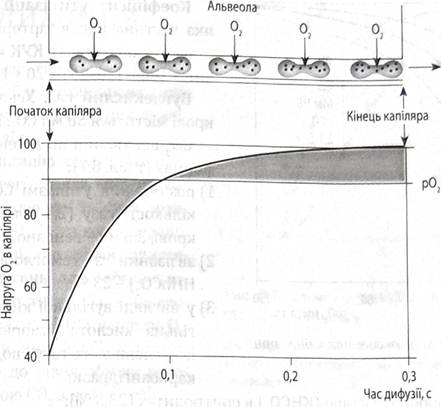 Рис. 8.2. Оксигенація крові в капілярах легень  Рис. 8.3. Крива дисоціації оксигемоглобіну. 132. Транспорт вуглекислий газу кров'ю. Роль еритроцитів у транспорті вуглекислий газу. Вуглекислий газ транспортується наступними шляхами: Розчинений у плазмі крові – близько 25 мл/л. Зв’язаний з гемоглобіном (карбгемоглобін) – 45 мл/л. У вигляді солей вугільної кислоти – букарбонати каліі та натрію плазми крові – 510 мл/л. Таким чином, у стані спокою кров транспортує 580 мл вуглекислого газу в 1 л. Отже, основною формою транспорту СО2 є бікорбонати плазми, що утворюються завдяки активному протіканню карбоангідразної реакції. В еритроцитах міститься фермент карбоангідраза (КГ), котрий каталізує взаємодію вуглекислого газу із водою з утворенням вугільної кислоти, що дисоціює з утворенням бікарбонатного йона та протона. Бікарбонат всередині еритроцита взаємодіє з йонами калію, що виділяються з калієвої солі гемоглобіну при відновленні останнього. Так всередині ер  итроцита утворюється бікарбонат калію. Так у плазмі утворюється бікарбонат натрію. итроцита утворюється бікарбонат калію. Так у плазмі утворюється бікарбонат натрію. В капілярах легень ці процеси йдуть в зворотньому напрямку. З йонів водню та бікарбонатних йонів утворюється вугільна кислота, котра швидко розпадається на вуглекислий газ та воду. Вуглекислий газ видаляється назовні. Отже, роль еритроцитів у транспорті вуглекислоти така: утворення солей вугільної кислоти; утворення карбгемоглобіну. 133. Фізіологічна роль діхальніх Шляхів, регуляція їх Просвіту. В дихальних шляхах повітря: - зігрівається; - зволожується, тому повітря в легенях насичене водяними парами на 100%, незалежно від вологості атмосферного повітря; - очищується, завдяки наявності війчастого епітелію та бокалоподібних клітин, які секретують слиз Окрім того, повітроносні шляхи мають велику кількість рецепторів (є рефлексогенною зоною), що забезпечує їх участь в здійсненні захисних дихальних рефлексів. В регуляції величини просвіту дихальних шляхів (і їх опору руху повітря) приймають участь, головним чином, нервові механізми. При цьому парасимпатичні рефлекторні впливи супроводжуються скороченням гладких м’язів Ù звуження дихальних шляхів. В гладких м’язах дихальних шляхів є a- та b-адренорецептори; стимуляція b-адренорецепторів супроводжується розслабленням гладких м’язів повітроносних шляхів і розширенням бронхіол, а стимуляція a-адренорецепторів – напруженям гладких м’язів та звуженням бронхів. В гладких м’язах дихальних шляхів переважають b-адренорецептори, тому в умовах підвищення активності симпато-адреналової системи просвіт дихальних шляхів розширюється Ù створюються оптимальні умови для вентиляції легень. 134. Діхальній центр, его будова, регуляція рітмічності дихання. Дихальний центр (ДЦ) знаходиться в задньому мозку (довгастий мозок та міст), хоча в регуляції зовнішнього жихання приймають участь всі рівні ЦНС, аж до кори головного мозку. Локалізація ДЦ визначається в дослідах з перерізкою стовбуру мозку. Таким чином, перерізка стовбуру вище заднього мозку не супроводжується змінами дихання, а перерізка нижче довгастого мозку призводить до його зупинки ДЦ знаходиться саме в задньому мозку. Дихальний центр довгастого мозку має два ядра – дорсальне та вентральне (розміщені симетрично по відношенню до середньої лінії). Особливістю дорсального ядра є наявність в ньому тільки інспіраторних нейронів, які збуджуються безпосередньо перед вдихом та під час вдиху (інспіраторні нейрони або нейрони вдиху). Нейрони дорсального дихального ядра забезпечують регуляцію дихання в стані спокою (при цьому акт вдиху є активним, а видиху – пасивним). Збудження інспіраторних нейронів дорсального ядра забезпечує скорочення м’язів спокійного вдиху вдих гальмування інспіраторних нейронів дорсального ядра розслаблення м’язів пасивний видих. До складу вентрального ядра входять як інспіраторні так і експіраторні нейрони (нейрони видиху). Це ядро вмикається в регуляцію форсованого (глибокого): дихання інспіраторні нейрони вентрального ядра забезпечують скорочення м’язів форсованого вдоху; експіраторні – м’язів видиху. Механізм ритмічного чергування вдиху та видихув стані спокою пов’язаний з ритмічним збдженням та гальмуванням інспіраторних нейронів дорсального ядра дихального центру. Зб  удження нейронів надходженням до них інформації від хеморецепторів (ХР) (центральних і периферичних), які є тонічно активними: генерують ПД. Збудження передається до мотонейронів спинного мозку збудження та скорочення дихальних м’язів вдих. удження нейронів надходженням до них інформації від хеморецепторів (ХР) (центральних і периферичних), які є тонічно активними: генерують ПД. Збудження передається до мотонейронів спинного мозку збудження та скорочення дихальних м’язів вдих.Далі збудження інспіраторних нейронів дорсального ядра змінюється їх гальмуванням. Цьому сприяє надходження до них інформації від таких структур: 1. Від рецепторів розтягнення легень (РРЛ) по аферентних волокнах блукаючого нерва спочатку до гальмівних вставних нейронів (ГВН) до інспіраторних нейронів; 2. Від пневмотаксичного центру (ПТЦ), збудження якого при вдосі пов’язане з надходженням до нього інформації від інспіраторних нейронів, а також від інших нейронів (ПТЦ – частина ретикулярної формації стовбура мозку і його аферентні зв’язки різноманітні). Коли потік гальмівної інформації від РРЛ та від ПТЦ до інспіраторних нейронів стає більш потужнішим, ніж потік збуджуючої інформації від хеморецепторів, інспіраторні нейрони гальмуються гальмування мотонейронів спинного мозку розслаблення дихальних м’язів видих. Видих знімає механізми, які активують РРЛ та ПТЦ припинення надходження гальмівної інформації до інспіраторних нейронів переважання аферентного входу від ХР збудження інспіраторних нейронів видих. 135. Механізм первого вдоху новонародженої дитини. 1. Після перерізки пуповини в крові дитини накопичується вуглекислота, знижується рН крові та знижується парціальний тиск кисню – стимуляція центральних та периферичних хеморецепторів – збудження дихального центру – збудження інспіраторних нейронів – скорочення м’язів вдиху. 2. Стимулювати перший вдих новонародженого можна дією на різні рецептори, оскільки аферентна інервація дихального центру дуже різноманітна: стимулюючи терморецептори – раніше дитину опускали то в холодну то в теплу воду; діючи на механорецептори (поплескування, пощипування шкіри); діючи на вестибулорецептори – дитину можна опускати головою вниз, нахиляти зі сторони в сторону. 3. Після народження дитини треба зняти “рефлекс пірнальника”, котрий блокує настання вдиху через наявність рідини в дихальних шляхах. Щоб зняти цей рефлекс цю рідину періодично аспірують. 136. Роль рецепторів розтягування легень и Блукаючи Нервів у регуляції дихання. Рецептори розтягнення легень збуджуються у відповідь на розтягнення легенів при вдосі, особливо при глибокому. Інформація від цих рецепторів передається в дихальний центр по аферентних волокнах блукаючого нерва і через гальмівні вставні нейрони гальмує інспіраторні нейрони видих. Таким чином, за участю цих рефлексів, які починаються з подразнення цих рецепторів при вдосі, вдих припиняється і наступає видих. Ці рефлекси носять назву рефлексів Герінга-Брайера. Вони приймають участь в саморегуляції ритму дихання. Цим і визначається роль блукаючих нервів в забезпеченні ритму дихання. в регуляції дихання відносяться: ірритантні рецептори легень, які реагують на дію їдких газів, пилу, тютюнового диму, холодного повітря і при збудженні зумовлюють звуження бронхів і гіпервентиляцію; юкстакапілярні рецептори, що розміщуються поблизу капілярів легень і реагують на зміну механічних властивостей легеневої тканини (набряк, переповнення капілярів кров’ю і т.д.) тахіпное; рецептори дихальних шляхів, що реагують на механічні та хімічні подразники і спричиняють виникнення захисних дихальних рефлексів (кашель, чхання, звуження бронхів). пропріорецептори працюючих м’язів,інформація від яких стимулює зовнішнє дихання (рефлекторно) при фізичному навантаженні; пропріорецептори дихальних м’язів– забезпечують рефлекторну саморегуляцію дихання при зміні опору (на вдиху чи на видиху). артеріальні пресорецептори, які реагують на підвищення САТ гіповентиляція. Протилежні реакції спостерігаються при зниженні САТ; вольові температурні рецептори– їх стимуляція призводить до різноманітних змін в акті дихання. 137. Роль центральних и периферичної хеморецепторів у регуляції дихання. Компоненти крови,что стімулюють Зовнішнє дихання. Периферичні хеморецептори локалізуються в судинних рефлексогенних зонах (дуга аорти і каротидні синуси). В регуляції дихання більш важливими є каротидні хеморецептори. Адекватні подразники для них: збільшення Рсо2 артеріальної крові; зменшення рН артеріальної крові; зменшення Ро2 артеріальної крові. Інформація, що надходить до дихального центру при підвищенні активності цих рецепторів, викликає гіпервентиляцію (підвищення глибини та частоти дихання) Центральні хеморецептори локалізуються на вентральній поверхні довгастого мозку (дно ІV шлуночка) і омиваються ліквором. Адекватними подразниками для цих рецепторів є: збільшення Рсо2 ліквора; зменшення рН ліквора. Підвищення активності цих рецепторів, як і периферичних, призводить до розвитку гіпервентиляції нормалізація вказаних показників ліквора. Таким чиномреагують на зниження Ро2 артеріальної крові. Тому гіпоксія стимулює зовнішнє дихання тільки за рахунок рефлексів, які розпочинаються з подразнення периферичних хеморецепторів. Таким чином, не дивлячись на велику роль в регуляції зовнішнього дихання центральних хеморецепторів, периферичні хеморецептори: забезпечують більш швидкі рефлекторні відповіді; тільки вони реагують на гіпоксію стимуляція зовнішнього дихання в умовах гіпоксії. До компонентів крові, які стимулюють зовнішнє дихання, відносяться Ро2, рН, Рсо2. Найбільшого значення для регуляції зовнішнього дихання має вміст в крові СО2 (Рсо2), менше – вміст йонів Н+, саме менше – вміст О2 (Ро2) При зниженні рН артеріальної крові нижче 7,4 вентиляція легень (ХОД) також підвищується. Залежність вентиляції легень від Рсо2 досліджують при диханні газовою сумішшю, в якій Ро2 залишається незмінним а Рсо2 поступово підвищується. Зниження Ро2 в артеріальній крові також супроводжується гіпервентиляцією. 138. Регуляція зовнішнього дихання при фізічному навантаженні. При фізичному навантаженні розвивається гіпервентиляція, ступінь якої пропорційна інтенсивності навантаження. Регуляція зовнішнього дихання в цих умовах здійснюється як по збуренню (ведучий механізм) так і по відхиленню (в умовах дуже інтенсивного або довготривалого навантаження). Головним механізмом розвитку гіпервентиляції при фізичному навантаженні є безумовні рефлекси з пропріорецепторів працюючих м’язів (керуючий пристрій (КП), яким є дихальний центр, отримує по каналу зовнішнього зв’язку інформацію від пропріорецепторів працюючих м’язів про роботу що виконується; КП аналізує цю інформацію і викликає підвищення глибини та частоти дихання для того, щоб при збільшених метаболічних потребах тканин склад альвеолярного повітря залишався незмінним). Рецептори з пропріорецепторів працюючих м’язів є ведучим, та не єдиним механізмом, який стимулює розвиток гіпервентиляції при фізичному навантаженні. Ще до початку роботи гіпервентиляція може розвиватися за механізмом умовних рефлексів (передстартові стани у спортсменів). Від моторної зони кори ГМ паралельно з передачею команди про початок роботи до скелетних м’язів, інформація надходить і до нейронів дихального центру, що також сприяє розвитку гіпервентиляції. |